Matrix genetics, part 2: the degeneracy of the genetic code and the octave algebra with two quasi-real units (the genetic octave Yin-Yang-algebra)
Algebraic properties of the genetic code are analyzed. The investigations of the genetic code on the basis of matrix approaches ("matrix genetics") are described. The degeneracy of the vertebrate mitochondria genetic code is reflected in the black-an…
Authors: Sergey V. Petoukhov (Department of Biomechanics, Mechanical Engineering Research Institute of the Russian Academy of Sciences)
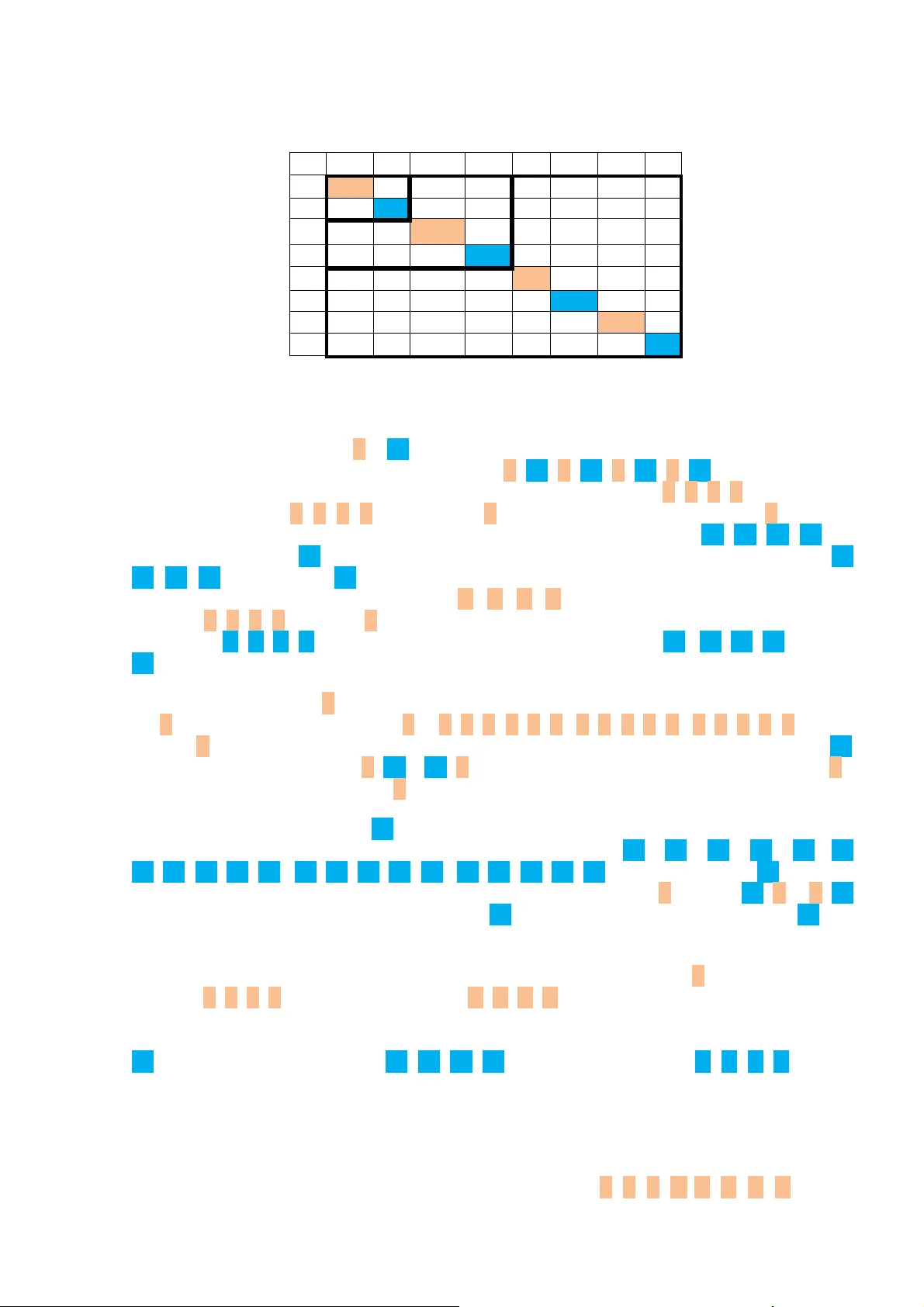
Matrix genetics, part 2: the degeneracy of th e genetic code and the octave algebra with two quasi-real units (the genetic octave Yin-Yang-algebra) Sergey V. Petoukhov Department of Biomechanics, Mechanical Engineering Research Institute of the Russian Academy of Sciences petoukhov@hotmail.com , petoukhov@im ash.ru , http://symm etry.hu/isabm/petoukhov.html Abstract . Algebraic properties of the genetic code are analyzed. The investigations of the genetic code on the basis of matrix approach es (“matrix genetics ”) are described. The degeneracy of the vertebrate mitochondria gene tic code is reflected in the black-and-white mosaic of the (8*8)-matrix of 64 triplets, 20 am ino acids and stop-signals. This m osaic genetic matrix is connected with the m atrix form of presentation of the specia l 8-dimensional Yin-Yang- algebra and of its particular 4-dimensional case. The special algorithm, which is based on features of genetic molecules, exists to tran sf orm the m osaic genomatrix in to the matrices of these algebras. Two new numeric systems are de fined by these 8-dim ensi onal and 4-dimensional algebras: genetic Yin-Yang-octave s and genetic tetrions. Their co mparison with quaternions by Hamilton is presented. Elements of new “genovector calcula tion” and ideas of “genetic mechanics” are discussed. These algebras are co nsid ered as models of the genetic code and as its possible pre-code basis. They are related with binary oppositions of the Yin-Yang type and they give new opportunities to investig ate evolution of the genetic code. The reve aled fact of the relation between the genetic code and these genetic algebras is discussed in connection with the idea by Pythagoras: ”All things are numbers”. Sim ultaneously these genetic algebras can be utilized as the algebras of genetic operators in biological organisms. Th e described results are related with the problem of algebraization of bioinform atics. Th ey take attention to the question: what is life from the viewpoint of algebra? KEYWORDS: genetic code, algebra, hypercompl ex num ber, quaternion, tetra-reproduction 1 Introduction This article is devoted to algebr aic properties of molecular system s of the genetic code in their matrix representations. T he initial approaches for investigations of genetic code systems from the viewpoint of matrix approach es were described in our pr evious publications [Petoukhov, 2001- 2008]. These investigations are generalized under the name “m atrix genetics”. They are connected closely with matrix forms of digital signal processing in computers. From an information viewpoint, biological organi sms are informational essences. They receive genetic information from their ancestors and transmit it to descendants. A conception of informational nature of living or ganisms is ref lected in the wo rds: “If you want to understand life, don’t think about vibrant, throbbing dels and oozes, thi nk about information technology” [Dawkins, 1991]. Or another ci tation of a similar direction of thoughts: “Notions of “information” or “valuable info rmation” are not utilized in phys ics of non-biological nature because they are not needed there. On the contrary, in biology notions “information” and especially “valuable information” are main one s; understanding and description of phenomena in biological nature are impossible without these noti ons. A specificity of “living matter” lies in them” [Chernavskiy, 2000]. Bioinformatics can give deeper knowledge in th e questions what is life and why life exists. A development of theor etical biology needs in appropriate m athematical models of structural ensembles of genetic elements. The effec tive matr ix approach for such models is proposed below. 2 The genetic octave matrix as the matrix form of presentation of the octave algebra Algebras of complex and hypercomplex numbers x 0 * 1 +x 1 * i 1 +…+x k * i k are well-known (the usual definition of the term “algebra over a field P” is given in the Appendix B). It is known also that complex and hypercomplex numbers have not only vector forms of their presentations, but also matrix forms of their presenta tion. For example com plex numbers z = x* 1 +y* i (where 1 is the real unit and i 2 = -1 is the imaginary unit) possess the fo llowing matrix form of their presentation: z = x* 1 + y* i = x* 1 0 0 1 + y* 0 1 -1 0 = x y -y x (1) By the way, complex numbers are utilized in comput ers in this m atrix form. The f ollowing table of multiplication of the basic matrix elements 1=[ 1 0; 0 1] and i = [ 0 1; -1 0] for the algebra of complex numbers exists: 1 i 11 i (2) ii - 1 Our initial idea is concluded in interpretation of the gen etic matrice s as matrix forms of presentation of special algebras ( or systems of m ultidimensional numbers) on the basis of molecular features of the letters C, A, U/T, G of the genetic alphabet. Let us apply this idea to the genetic matrix P CAUG 123 (3) (or P (3) =[C A; U G] (3) ), which was described in our previous articles [Petoukhov, arXiv:0802.3366;e of the vertebrate mitochondria genetic code (Figure 1). This geno matrix has 32 “black” triplets and 32 “white” triplets disposed in matrix cells of a ppropriate colors (see details in [Petoukhov, genomatrix possess identical mosaics. All triple ts in two quadrants along the m ain diagonal begin with the letters C and G, which possess 3 hydrogen bonds in their complementary pair C-G. All triplets in two quadrants along the se cond diagonal begin with the letters A and U, which possess 2 hydrogen bonds in their complementary pair A-U. The phenomenological “alphabetic” rule # 1 exists for the matrix d isposition of triplets of black and white colors: - In the set of 32 triplets, the first letter of which has 3 hydr ogen bonds (the letters C and G), the white triplets are those one s, the s econd position of which is occupied by the letter A (that is by the purin e with 2 hydrogen bonds); the othe r triplets of this set are black triplets; - In the set of 32 triplets, the first letter of which has 2 hydrogen bonds (the letters A and U), the black triplets are those ones, the sec ond position of which is occupied by the letter C (that is by the pyrimidine with 3 hydrogen bonds ); the othe r triplets of this set a re white triplets. For example in accordance with this rule th e trip let CAG is the white trip let because its first letter C has 3 hydrogen bonds, and its second positio n is occupied by the letter A. It is obvious that the first part of this rule, which utilizes mol ecula r features of the genetic le tters C, A, G, U, is related to tr iplets of two quadrants along the main matr ix diagonal, and that the second part of this rule is related to triplets of two quadrants along the second diagonal. We should note the inessential modification in nu meration of the columns and the rows in this article in comparison with our previ decreasing sequence 111 (7), 110 (6 ), 101 (5), 100 (4), 011 (3), 010 (2), 001 (1), 000 (0) in the genomatrix P CAUG 123 (3) was utilized. It was made to demonstr ate the coincidence with the fam ous table of 64 hexagrams of the ancient Chinese book “I Ching”, which po ssessed this decreasing sequence. Now we number the columns and th e rows of this genomatrix on Figure 1 by the ascending sequence 000 (0), 001 (1), 010 (2), 011 (3), 100 (4), 101 (5), 110 (6), 111 (7), which is more traditional for matrix analysis and fo r the theory of digital signal processing. The columns and the rows are numbered by this asce nding sequence on the basis of their triplets algorithmically, if we change the correspondence between binary symbols (0 and 1) and the genetic letters in the two first genetic s ub-alphabets [Petoukhov, arXiv:0803.0888] by assum ing the following: P CAUG 123 (3) : 000 (0) 001 (1) 010 (2) 011 (3) 100 ( 4) 101 (5) 11 0 (6) 111 (7) 000 (0) CCC Pro CCA Pro CAC His CAA Gln ACC Thr ACA Thr AAC Asn AAA Lys 001 (1) CCU Pro CCG Pro CAU His CAG Gln ACU Thr ACG Thr AAU Asn AAG Lys 010 (2) CUC Leu CUA Leu CGC Arg CGA Arg AUC Ile AUA Met AGC Ser AGA Stop 011 (3) CUU Leu CUG Leu CGU Arg CGG Arg AUU Ile AUG Met AGU Ser AGG Stop 100 (4) UCC Ser UCA Ser UAC Tyr UAA Stop GCC Ala GCA Ala GAC Asp GAA Glu 101 (5) UCU Ser UCG Ser UAU Tyr UAG Stop GCU Ala GCG Ala GAU Asp GAG Glu 110 (6) UUC Phe UUA Leu UGC Cys UGA Trp GUC Val GUA Val GGC Gly GGA Gly 111 (7) UUU Phe UUG Leu UGU Cys UGG Trp GUU Val GUG Val GGU Gly GGG Gly Figure 1. The genomatrix P CAUG 123 (3) , each cell of which has a triplet and an amino acid (or stop - signal) coded by this triplet. The black-and-white mosaic presents a specificity of the degeneracy of the vertebrate mitochondria gene 000 (0) 001 (1) 010 (2) 011 (3) 100 (4) 101 (5) 110 (6) 111 (7) 000 (0) x 0 x 1 -x 2 -x 3 x 4 x 5 -x 6 -x 7 001 (1) x 0 x 1 -x 2 -x 3 x 4 x 5 -x 6 -x 7 010 (2) x 2 x 3 x 0 x 1 -x 6 -x 7 -x 4 -x 5 YY 8 = 011 (3) x 2 x 3 x 0 x 1 -x 6 -x 7 -x 4 -x 5 100 (4) x 4 x 5 -x 6 -x 7 x 0 x 1 -x 2 -x 3 101 (5) x 4 x 5 -x 6 -x 7 x 0 x 1 -x 2 -x 3 110 (6) -x 6 -x 7 -x 4 -x 5 x 2 x 3 x 0 x 1 111 (7) -x 6 -x 7 -x 4 -x 5 x 2 x 3 x 0 x 1 Figure 2. The matrix YY 8 as the matrix form of presentation of the genetic octave algebra with two quasi-real units (the geneti c octave Yin-Yang-algebra). The black cells contain coordinates with the sign „+” and the white ce lls c ontain coordinates with the sign „-”. • the first genetic sub-alphabet, that defines the binary number of the columns, presents each pyrimidine (C and U/T) by the sym bol 0, and presents each purine (A and G) by the symbol 1; • the second genetic sub-alphabet, that defines the binary nu mber of the rows, presents each letter with the amino-mutatin g pr operty (C and A) by the symbol 0 , and presents each letter without such propert y (G and U/T) by the symbol 1 . Taking into account the molecular characteristics of the nitrogenous bases C, A, U/T, G of the genetic alphabet, one can reform the genomatrix P CAUG 123 (3) into the new matrix YY 8 algorithmically (Figure 2). The m osaic of the di sposition of signs “+” (it occupies the black matrix cells) and “-“ (it occupies th e white matrix cells) in matr ix YY 8 is identical to the m osaic of the genomatrix P CAUG 123 (3) (Figure 1). Below we shall list the stru ctural analogies be tween these matrices P CAUG 123 (3) and YY 8 and demonstrate that this matrix YY 8 is the matrix representation of the octave algebra with two qua si-real units. But initially we pay attention to the “alphabetic” a lgorithm of digitization of 64 triplets, which giv es the matrix YY 8 from the genomatrix P CAUG 123 (3) . 2.1 The alphabetic algorithm of the Yi n-Yang-digitization of 64 triplets This algorithm is based on utilizing the two binary -oppositional attributes of the genetic letters C, A, U/T, G: “purine or pyrimidine” and “2 or 3” hydrogen bonds. It uses also the fam ous thesis of molecular genetics that different positions insi de triplets have different code meanings. For example the article [Konopelchenko, Rumer, 1975] ha s described that two first positions of each triplet form “the root of the c odon” and that they differ drastically from the third position by their essence and by their special role. Because of th is “alphabetic” algorithm, the transfo rmation of the genomatrix P CAUG 123 (3) into the matrix YY 8 is not an abstract and ar bitrary action at all, but such transformation can be utilized by bioc omputer system s of or ganisms practically. The alphabetic algorith m of the Yin-Yang-digitization defines the special scheme of reading of each triplet: the first two positions of the triplet are read by ge netic sys tems from the viewpoint of one attribute (the attribute of “2 or 3” hyd rogen bonds) and the third pos ition of the triplet is read from the viewpoint of anot her attribute (the attribute of “purine or pyrim idine”). The algorithm consists of three parts, where the f irst two parts define the generalized num eric symbol of each triplet and the third part defines its sign “+” or “-“: 1. Two first positions of each triplet is read fr om the viewpoint of the binary-opposition al attribute “2 or 3 hydrogen bonds” of the ge netic letters: each letter from the complementary pair C and G is interpreted as a real number α (for instance, α =3 because C and G have 3 hydrogen bonds each), and each letter from the second complementary pair A and U/T is interpreted as a real number β (for instance, β =2 because A and U/T have 2 hydrogen bonds each). 2. The third position of each triplet is read from the viewpoint of the another binary- oppositional attribute “purine or pyrim idine”: each pyrimidine C or U/T is interpreted as a real number γ (for instance γ =1 because each pyrim idine c ontains one m olecular ring), and each purine A or G is interpreted as a real num ber δ (for instance, δ =2 because each purine contains two molecular rings). 3. The generalized numeric symbol of each black (white ) triplet has a sign “+” (“-“ correspondingly); the definition of the bl ack triplets and the wh ite triplets was made above in the rule # 1 on the basis of molecu lar properties of the ge netic letters also. For example the triplet CAG receives the generalized num eric symbol “- αβδ ” by this algorithm because its first letter C is symbolized as “ α ”, the second letter is sym bolized as “ β ” and the third letter G is symbolized as “ δ ”. The sign “-“ appears because CAG is the white tr iplet in accordance with the “alphabetic” rule # 1. The desc ribed algorithm can be considered as the algorithm of special con version by means of which the fo ur genetic letters are substitu ted for four real numbers α , β , γ , δ , and each triplet appears in the form of the chain (or the ensemble) of these real numbers with the appropriate sign. One can say that new alphabet of the four symbols α , β , γ , δ appears (see the section 6 about the numeric sy stem of tetrions as the genetic pre-code). Figure 3 illustrates the detail s of such algorithmic conversion of the genomatrix P CAUG 123 (3) into the matrix YY 8 , where the 8 variants of the 3-digit chai ns take place as co mponents of the matrix YY 8 : ααγ =x 0 , ααδ =x 1 , αβγ =x 2 , αβδ =x 3 , βαγ =x 4 , βαδ =x 5 , ββγ =x 6 , ββδ =x 7 . We shall name these matrix components x 0 , x 1 ,…, x 7 , which are real numbers, as “YY-coordinates”. 000 (0) 001 (1) 010 (2) 011 (3) 100 (4) 101 (5) 110 (6) 111 (7) 000 (0) CCC ααγ x 0 CCA ααδ x 1 CAC - αβγ -x 2 CAA - αβδ -x 3 ACC βαγ x 4 ACA βαδ x 5 AAC - ββγ -x 6 AAA - ββδ -x 7 001 (1) CCU ααγ x 0 CCG ααδ x 1 CAU - αβγ -x 2 CAG - αβδ -x 3 ACU βαγ x 4 ACG βαδ x 5 AAU - ββγ -x 6 AAG - ββδ -x 7 010 (2) CUC αβγ x 2 CUA αβδ x 3 CGC ααγ x 0 CGA ααδ x 1 AUC - ββγ -x 6 AUA - ββδ -x 7 AGC - βαγ -x 4 AGA - βαδ -x 5 011 (3) CUU αβγ x 2 CUG αβδ x 3 CGU ααγ x 0 CGG ααδ x 1 AUU - ββγ -x 6 AUG - ββδ -x 7 AGU - βαγ -x 4 AGG - βαδ -x 5 100 (4) UCC βαγ x 4 UCA βαδ x 5 UAC - ββγ -x 6 UAA - ββδ -x 7 GCC ααγ x 0 GCA ααδ x 1 GAC - αβγ -x 2 GAA - αβδ -x 3 101 (5) UCU βαγ x 4 UCG βαδ x 5 UAU - ββγ -x 6 UAG - ββδ -x 7 GCU ααγ x 0 GCG ααδ x 1 GAU - αβγ -x 2 GAG - αβδ -x 3 110 (6) UUC - ββγ -x 6 UUA - ββδ -x 7 UGC - βαγ -x 4 UGA - βαδ -x 5 GUC αβγ x 2 GUA αβδ x 3 GGC ααγ x 0 GG А ααδ x 1 111 (7) UUU - ββγ -x 6 UUG - ββδ -x 7 UGU - βαγ -x 4 UGG - βαδ -x 5 GUU αβγ x 2 GUG αβδ x 3 GGU ααγ x 0 GGG ααδ x 1 Figure 3. The result of the algorithmic conferment of 64 triplets to num eric coordinates x 0 , x 1 , …, x 7 , which are based on the four real numbers α , β , γ , δ . In the section 2.3 we will describe the structural analogies between sets of elements of the genomatrix P CAUG 123 (3) and of the matrix YY 8 . But now we will pay attention to algebraic properties of the matrix YY 8 . 2.2 The Yin-Yang-genomatrix YY 8 as the element of the octave Yin-Yang-algebra It is quite unexpectedly that this new m atrix YY 8 (Figure 2), which is constructed algorithm ically from the genom atrix P CAUG 123 (3) , presents the unusual algebra w ith such set of basic elements which contains two quasi-real units and does not c ontain the real unit at all. Really the matrix YY 8 with its 8 coordinates x 0 , x 1 ,…, x 7 can be represented as the sum of the 8 m atrices, each of which contains only one of these coordinates (Figure 4). Let us symbolize any matrix, which is multiplied there by any of YY-coordinates x 0 , x 2 , x 4 , x 6 with even indexes, by the symbol f k (where “f” is the first letter of the word “fem ale” and k=0, 2, 4, 6). We will m ark these matrices f k and their coordinates x 0 , x 2 , x 4 , x 6 by pink color. And let us sy mbolize any matrix, which is multiplied there by any of YY-coordinates x 1 , x 3 , x 5 , x 7 with odd indexes, by the symbol m p (where “m” is the firs t letter of the word “m ale” and p=1, 3, 5, 7). W e will mark th ese matrices m p and their coordinates x 1 , x 3 , x 5 , x 7 by blue color. In this case one can present the matrix YY 8 in the form (3). YY 8 = x 0 * f 0 +x 1 * m 1 +x 2 * f 2 +x 3 * m 3 +x 4 * f 4 +x 5 * m 5 +x 6 * f 6 +x 7 * m 7 (3) YY 8 = х 0 * 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 + х 1 * 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 + + х 2 * 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 + х 3 * 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 + + х 4 * 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 + х 5 * 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 + + х 6 * 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 + х 7 * 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 -1 0 0 0 0 0 0 0 -1 0 0 0 0 0 0 Figure 4. The presentation of the matrix YY 8 as the sum of the 8 m atrices . The important fact is that the set of these 8 matrices f 0 , m 1 , f 2 , m 3 , f 4 , m 5 , f 6 , m 7 forms the closed set relative to m ultiplications: a multiplication be tw een any two matrices from this set gener ates a matrix from this set again. The table on Figure 5 presents the resu lts of mu ltiplications among these 8 matrices. It is known that s uch multipli cation table s define appropriate alg ebras over a field (see Appendix B). Correspondingly the table on Figure 5 defines the genetic octave algebra YY 8 . Multiplication of any two m embers of the octave algeb ra YY 8 generates a new member of the same algebra. This situation is similar to the situatio n of real num bers (or of com plex numbers, or of hypercomplex numbers) when m ul tiplication of any two members of the numeric system generates a new member of the sam e numerical system. In other words, we receive new numerical system of YY 8 octaves (3) from the natural structure of the genetic code. f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 f 0 f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 m 1 f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 f 2 f 2 m 3 - f 0 - m 1 - f 6 - m 7 f 4 m 5 m 3 f 2 m 3 - f 0 - m 1 - f 6 - m 7 f 4 m 5 f 4 f 4 m 5 f 6 m 7 f 0 m 1 f 2 m 3 m 5 f 4 m 5 f 6 m 7 f 0 m 1 f 2 m 3 f 6 f 6 m 7 - f 4 - m 5 - f 2 - m 3 f 0 m 1 m 7 f 6 m 7 - f 4 - m 5 - f 2 - m 3 f 0 m 1 Figure 5. The multiplica tion table of the Yin-Yang-algebra Y Y 8 for the case of P CAUG 123 (3) . In accordance with its multiplicatio n table (Figure 5), th e algebra of this new numeric system contains two quasi-real units f 0 и m 1 in the set of its 8 basic matrices and it does not contain the real unit 1 at all. Really the set of basic matrices f 0 , m 1 , f 2 , m 3 , f 4 , m 5 , f 6 , m 7 is divided into two equal sub-sets by attributes of their squares. The first sub-set contains f 0 , f 2 , f 4 , f 6 .The squares of these basic matrices f 0 , f 2 , f 4 , f 6 are equal to ± f 0 always. We name this sub-set the f 0 -sub-set of the basic matrices (or of the basic el ements). The second sub-set contains m 1 , m 3 , m 5 , m 7 . We name this sub-set the m 1 -sub-set of the basic elements. The squares o f these basic matrices m 1 , m 3 , m 5 , m 7 are equal to ± m 1 always (see elements on th e ma in diagonal of the multiplication table on Figure 5). The YY-coordinates x 0 , x 2 , x 4 , x 6 , which are connected with the basic elements f 0 , f 2 , f 4 , f 6 , form the f 0 -sub-set of the eight YY-coor dinates correspondingly. The YY- coordinates x 1 , x 3 , x 5 , x 7 , which are connected with the basic elements m 1 , m 3 , m 5 , m 7 , form the m 1 -sub-set. The initial basic element f 0 possesses all properties of the real unit in relation to each member of the f 0 -sub-set of the basic elem ents: f 0 2 = f 0 , f 0 * f 2 = f 2 * f 0 = f 2 , f 0 * f 4 = f 4 * f 0 = f 4 , f 0 * f 6 = f 6 * f 0 = f 6 . But the element f 0 loses the commutative property of the real un it in relation to the memb ers of the m 1 - sub-set of the basic elements: f 0 * m p ≠ m p * f 0 , where p = 1, 3, 5, 7. In this reason the element f 0 is named the quasi-real unit from the f 0 -sub-set of the basic elements. By analogy the basic element m 1 possesses all properties of the real unit in relation to each membe r of the second sub-set of the basic elements m 1 , m 3 , m 5 , m 7 : m 1 2 = m 1 , m 1 * m 3 = m 3 * m 1 = m 3 , m 1 * m 5 = m 5 * m 1 = m 5 , m 1 * m 7 = m 7 * m 1 = m 7 . But the element m 1 loses the commutative property of the real unit in re lation to members of the f 0 -sub-set: m 1 * f k ≠ f k * m 1 , where k = 0, 2, 4, 6. In this reason the element m 1 is named the quasi-real unit from the m 1 -sub- set of the basic elements. Let us pay attention to the unexpected circumstance. All m embers of the f 0 -sub-set of the basic elements f 0 , f 2 , f 4 , f 6 and of their coordinates x 0 , x 2 , x 4 , x 6 have the even indexe s 0, 2, 4, 6 (zero is considered as even number here). And they o ccupy the columns with the even numbers 0, 2, 4, 6 in the YY 8 -matrix (Figure 2) and in the multiplication table (Figure 5). All members of the m 1 -sub-set of the basic elements m 1 , m 3 , m 5 , m 7 and of their coordinates x 1 , x 3 , x 5 , x 7 have the odd indexes 1, 3, 5, 7. And they occupy the columns with the odd numbers 1, 3, 5, 7 in the YY 8 - matrix (Figure 2) and in the m ultiplication table (Figure 5). By Pythagorean and Ancient Chinese traditions all even nu mbers are named “fem ale” numbers or Yin-numbers, and all odd numbers are named “male” numbers or Yang-numbers. In accordance with these traditions on e can name the elem ents f 0 , f 2 , f 4 , f 6 , х 0 , х 2 , х 4 , х 6 with the even indexes as “female” or Yin-elem ents and the elements m 1 , m 3 , m 5 , m 7 , x 1 , x 3 , x 5 , x 7 with the odd indexes as “male” elements or Yang-elem ents conditionally. By analogy one can name the columns with the even numerations 0, 2, 4, 6 (with the odd num erations 1, 3, 5, 7) as the female columns (the male columns). In this r eason this octave algebra of the genetic code is named “the octave algebra with two quasi-real units” or the octave Yin-Yang-algebra (or the bisex algebra, or the even-odd-al gebra). Such algebra gives new e ffective possibilities to model binary oppositions in biological objects at different levels, including triplets, amino acids, m ale and female gametal cells, m ale and female chromosomes, etc. Each genetic triplet, which is disposed toge ther with one of the female YY-coordin ates x 0 , x 2 , x 4 , x 6 in a mutual matrix c ell, is named the fem ale triplet o r the Yin-triplet (Figure 3 ). The third position of all female triplets is occu pied by the letter γ , which corresponds to the pyrim idine C or U/T. In this reason the female triplets can be n amed “pyrim idine triplets” as well. Each triplet, which is disposed together with one of the male YY-coordinates x 1 , x 3 , x 5 , x 7 in a mutual matrix cell, is named the male triple t or the Yang-triplet. The third pos ition of all m ale triplets is occupied by the letter δ , which corresponds to the purine A or G. In this reason the male triplets can be named “purine triplets”. In su ch algebraic way the whole set of 64 triplets is divided in to two sub-sets of Yin-triplets (or fem ale triplets) and Yang-triplets (or m ale triplets). We shall demonstrate later that the set of 20 amino acids is divided into the su b-sets of “fem ale amino acids”, “male amino acids” and “androgyne am ino acids” from the this matrix viewpoint. The multiplication table (Figure 5 ) is not symme tric one relativ e to the main diagonal; it corresponds to the non -commutative property of the Yin-Yang al gebra. The expression (3) is the vector form of presentation of the genetic YY 8 -number for the case of the genomatrix P CAUG 123 (3) . It reminds the vector form of pr esentation of hypercom plex numbers x 0 * 1 +x 1 * i 1 +x 2 * i 2 +x 3 * i 3 +x 4 * i 4 +… . But the significant d ifferen ce exists between hypercomplex numbers and Yin-Yang-num bers. All cells of th e main diagonal of multiplicatio n tables for hypercomplex numbers are occupied always by the real unit only (with the signs “+” or “-“). On the contrary, all cells of the main di agonal of multiplication tables for YY 8 -numbers are occupied by two quasi-real units f 0 and m 1 (with the signs “+” or “-“) without the real unit at all (Figure 5). By their definition “hypercomplex numbers are the elem ents of the algebras with th e real unit” [Mathematical encyclopedia, 1977]. Complex a nd hypercom plex numbers were constructed historically as generali zations of real numbers with the obligatory in clus ion of the real unit in sets of their basic elements. It can be dem onstrat ed easily that Yin-Yang algebras are the specia l generalization of the algebras of hypercomplex num bers. YY-numbers becom e the appropriate hypercomplex numbers in those cases when all th eir female (or male) c oordinates are equal to zero. In other words, YY-numbers are the specia l generalization of hypercomplex numbers in the form of “double-hypercomplex” numbers. Traditi onal hypercomplex numbers can be represented as the “mono-sex” (Yin or Yang) half of appropriate YY-num b ers. The algorithm of such generalization will be describe d later. We will denote Yin-Y a ng numbers by double letters (for example, YY) to distinguish th em from tradit ional (complex and hypercom plex) numbers. In comparison with hypercomplex numbers, Yin-Y ang numbers are the new ca tegory of numbers in the mathematical natural sciences in principl e. In our opinion, knowledge of this category of numbers is necessary for deep unde rstanding of biological phenom en a, and, perhaps, it will be useful for mathem atical natural sciences in the whole. Mathematical theory of YY-num bers gives new formal and conceptual apparatus to m odel phenomena of reproduction, self- organization and self-development in living nature. The set of the basic elements of the YY 8 -algebra forms a semi-group. Two squares are marked out by bold lines in the left upper corner of the multiplication table on Figure 5. The first two basic elements f 0 and m 1 are disposed in the smaller (2x2)-s quare of this table only. The greater (4x4)-square collects the four first basic elements f 0 , m 1 , f 2 , m 3 , which do not meet outside this square in the table also. These aspects say that sub-algebras YY 2 and YY 4 exist inside the algebra YY 8 . We shall return to these sub-algebras in the Appendix A.2. 2.3 The structural analogies between the genomatrix P CAUG 123 (3) and the matrix YY 8 One should remind that the black cells of the genomatrix P CAUG 123 (3) contain the black NN- triplets, which encode the 8 high-degeneracy am ino acids and the coding meaning of which do not depend on the letter on their thir d position (see details in [Petoukhov, arXiv:0802.3366 ; arXiv:0803.0888 ]). The set of the 8 high-degeneracy amino acids contains those amino acids, each of which is encoded by 4 triplets or more: Ala, Arg, Gly, Leu, Pro, Ser, Thr, Val [Petoukhov, 2005]. The white cells of the genomatrix P CAUG 123 (3) contain the white NN-triplets, the coding meaning of which depends on th e letter on th eir thir d position and which encode the 12 low- degeneracy amino acids together with stop-signal s. And the set of the 12 low-degeneracy amino acids contains those amino acids, each of which is encoded by 3 triplets or less: Asn, Asp, Cys, Gln, Glu, His, Ile, Lys, Met, Phe, Trp, Tyr. The table on Figure 6 shows signi ficant analogies and interrel ations between the matrix YY 8 and the genomatrix P CAUG 123 (3) (Figures 1-3). Such structural coincidence of two matrices YY 8 and P CAUG 123 (3) permits to consider the octave algebr a YY 8 as the adequate mode l of the structure of the genetic code. One can postulate such algebr aic model and then deduce som e peculiarities of the genetic code from this model. The octave Yin-Yang matrix YY 8 The octave genomatrix P CAUG 123 (3) This matrix possesses the binary m osaic of symmetrical character. It contains 32 YY-coordinates with the sign “+” and 32 YY-coordinates with the sign “-“. This genomatrix possesses the same binary mosaic. The black cells contain the high- degeneracy amino acids which are encoded by the 32 black NN-triplets. The white cells contain th e low-degeneracy amino ac ids and the stop-signals which are encoded by the 32 white NN- triplets. The enumerated matrix rows 0 and 1, 2 and 3, 4 and 5, 6 and 7 are equivalent to each other by a disposition of identical YY-coordinates. The enumerated matrix rows 0 and 1, 2 and 3, 4 and 5, 6 and 7 are equivale nt to each other by a disposition of identical amino acids. The half of kinds of YY-coordinates (x 0 , x 1 , x 2 , x 3 ) is presented in the quadrants along the main matrix diagonal only. The second half of kinds of YY-coordinates (x 4 , x 5 , x 6 , x 7 ) is presented in the quadrants along the second coordinates only. The half of kinds of amino acids is presented in the quadrants along the ma in matrix diagonal only (Ala, Arg, Asp, Gln, Glu, Gly, His, Leu, Pro, Val). The second half of kinds of amino acids is presented in the quadrants along the second diagonal only (Asn, Cys, Ile, Lys, Met, Phe, Ser,Thr, Trp, Tyr). The YY-coordinates x 0 , x 2 , x 4 , x 6 from the f 0 -sub-set occupy the columns with even numbers 0, 2, 4, 6. The YY- coordinates x 1 , x 3 , x 5 , x 7 from the m 1 - sub-set occupy the co lumns with the odd numbers 1, 3, 5, 7. The triplets with the pyrim idine C or U/T on their third position occupy the columns with the even numbers 0, 2, 4, 6. The trip lets with the purine A or G on their third position occupy the columns with the odd numbers 1, 3, 5, 7. Figure 6. Examples of the conf orm ity between the matrix YY 8 and the genomatrix P CAUG 123 (3) . The results of the comparison analysis in this table give th e following answer to the question about mysterious principles of the degeneracy of the genetic code from the viewpoint of the proposed algebraic model. The matrix dispos ition of the 20 amino acids and the sto p-signals is determined in some essential features by algeb rai c principles of the matrix disposition of the YY-coordinates. Moreover the disposition of the 32 black triplets and the high-degeneracy amino acids is determined by the disposition of the YY-coordinates with the sign “+”; the disposition of the 32 white triple ts, the low- degeneracy amino acids and stop-signals is determined by the disposition of the YY-coordinate s with th e sign “-”. One can rem ind here that the division of the set of the 20 amino acids into the two sub-sets of the 8 high-degeneracy amino acids and the 12 low-degeneracy amino acids is the i nvariant rule of all the dialects of the genetic code practically (see details in [Petoukhov, 2005]). The data of the table on Figure 6 do not exhaust the interconnections betw een the genetic code systems a nd the Yin-Yang matrices at all [Petoukhov, 2008b]. 3 The Yin-Yang octave algebras and the p ermutations of positions in triplets Our previous article [Petoukhov, arXiv:0803.0888] has presented the fact that the six possible variants of permutation s of three position s in triplets (1-2-3, 2-3-1, 3-1-2, 1-3-2, 2-1-3, 3-2-1 ) generate the family of the six genom atrices P CAUG 123 (3) , P CAUG 231 (3) , P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) . For the considered case of the vertebrate mitochondria genetic code, all these genomatrices have symmetrologic m osaics of the code degeneracy. These data says that the degeneracy of the code has non- trivial connections with the posit ion permutations in triplets. Each triplet has its own YY- coordinate from the set x 0 , x 1 , …, x 7 with the appropria te sign (Figure 3). The position permutatio ns in triplets l eads to new m atrix dispositions of the triplets together with their coordinates x 0 , x 1 , …, x 7 . In such way new genomatrices P CAUG 231 (3) , P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) originate from the initial genom atrix P CAUG 123 (3) . Algebraic properties of these matric es can be analyzed specially. Above we have demonstrated that the initial genomatrix P CAUG 123 (3) of this permutation family possessed the interrelation with the YY 8 -algebra. But the described permutation transformation of the genomatrix P CAUG 123 (3) into new genomatrices P CAUG 231 (3) , P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) can destroy this interrelation. For ex ample the set of basic elem ents of each of these new genomatrices can be an unclosed set, and algebras do not originate in this cas e, or this set can be connected wi th algebras of quite other type . One can demonstrate in g eneral case that arbitrary perm ut ations of the columns a nd of the rows of the P CAUG 231 (3) lead in general case to new matrice s, which possess unclosed sets of their b asic elements. For insta nce, if the first and the second columns in th e matrix P CAUG 231 (3) (or in the matrix YY 8 on Figure 2) interchange their places, the new matrix does not fit the YY-algebr a at all. Generally speaking, a very little probability e xists that these new ge nomatrices P CAUG 231 (3) , P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) fit YY 8 -algebras also. B ut if this unexpect ed fact would be revealed, this fact will be the strong evidence of the deep interrelation b etween Yin-Yang algebras and the genetic code additionally. Such unexpected fact was revealed by the au thor really: each of the genom atrices P CAUG 231 (3) , P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) fits their own YY 8 -algebra. Each of these genomatrices (with th e eight coordinates x 0 , x 1 , …, x 7 , which correspo nd to proper triplets ) possesses its own set of the eight basic elem ents and its own multiplication table, which determines an octave algebra with two quasi-r eal units also. W e shall mark these Yin-Yang algebras by the symbols (YY 8 ) CAUG 231 , (YY 8 ) CAUG 312 , etc. by analogy with the appropriate genomatrices P CAUG 231 (3) , P CAUG 312 (3) , etc. Each of these algebras possesses its own set of basic elements f 0 , m 1 , f 2 , m 3 , f 4 m 5 , f 6 , m 7 . In other words, the matrix presentations of these basic elements differ from each other in the cases of the different algebras (YY 8 ) CAUG 231 , (YY 8 ) CAUG 312 , etc., though we utilize the same symbols for them here. And both quasi-real units have different forms of their matrix presenta tions in different Yin-Yang octa ve algebras also. Figure 7 shows the example of the genomatrix P CAUG 231 (3) with the same coordinate from the set x 0 , x 1 , …, x 7 for each triplet as in the genomatrix P CAUG 123 (3) on Figure 3. It can be checked that the (8*8)-matrix with such disposition of coordinates x 0 , x 1 , …, x 7 is the matrix form of presentation of the Yin-Yang octave algebra (YY 8 ) CAUG 231 , the multiplication table of which is shown on Figure 8. The basic elements f 0 and m 4 occupy the main diagonal and play the role of the quasi-real units for this algebra. Figure 7. The disposi tion of coordinates x 0 , x 1 , …, x 7 in the genomatrix P CAUG 231 (3) reproduced from the artic CCC x 0 CAC -x 2 ACC x 4 AAC -x 6 CCA x 1 CAA -x 3 ACA x 5 AAA -x 7 CUC x 2 CGC x 0 AUC -x 6 AGC -x 4 CUA x 3 CGA x 1 AUA -x 7 AGA -x 5 UCC x 4 UAC -x 6 GCC x 0 GAC -x 2 UCA x 5 UAA -x 7 GCA x 1 GAA -x 3 UUC -x 6 UGC -x 4 GUC x 2 GGC x 0 UUA -x 7 UGA -x 5 GUA x 3 GGA x 1 CCU x 0 CAU -x 2 ACU x 4 AAU -x 6 CCG x 1 CAG -x 3 ACG x 5 AAG -x 7 CUU x 2 CGU x 0 AUU -x 6 AGU -x 4 CUG x 3 CGG x 1 AUG -x 7 AGG -x 5 UCU x 4 UAU -x 6 GCU x 0 GAU -x 2 UCG x 5 UAG -x 7 GCG x 1 GAG -x 3 UUU -x 6 UGU -x 4 GUU x 2 GGU x 0 UUG -x 7 UGG -x 5 GUG x 3 GGG x 1 f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 f 0 f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 f 1 f 1 - f 0 - f 3 f 2 m 5 - m 4 - m 7 m 6 f 2 f 2 f 3 f 0 f 1 m 6 m 7 m 4 m 5 f 3 f 3 - f 2 - f 1 f 0 m 7 - m 6 - m 5 m 4 m 4 f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 m 5 f 1 - f 0 - f 3 f 2 m 5 - m 4 - m 7 m 6 m 6 f 2 f 3 f 0 f 1 m 6 m 7 m 4 m 5 m 7 f 3 - f 2 - f 1 f 0 m 7 - m 6 - m 5 m 4 Figure 8. The multiplica tion table of the basic elem ents of the octave Yin-Yang-algeb ra (YY 8 ) CAUG 231 for the genomatrix P CAUG 231 (3) on Figure 7. The elements f 0 and m 4 are the quasi-real units in this alg ebra. f 0 f 1 m 2 m 3 f 4 f 5 m 6 m 7 f 0 f 0 f 1 m 2 m 3 f 4 f 5 m 6 m 7 f 1 f 1 f 0 m 3 m 2 f 5 f 4 m 7 m 6 m 2 f 0 f 1 m 2 m 3 f 4 f 5 m 6 m 7 m 3 f 1 f 0 m 3 m 2 f 5 f 4 m 7 m 6 f 4 f 4 - f 5 m 6 - m 7 - f 0 f 1 - m 2 m 3 f 5 f 5 - f 4 m 7 - m 6 - f 1 f 0 - m 3 m 2 m 6 f 4 - f 5 m 6 - m 7 - f 0 f 1 - m 2 m 3 m 7 f 5 - f 4 m 7 - m 6 - f 1 f 0 - m 3 m 2 Figure 9. The multiplica tion table of the basic elem ents of the octave Yin-Yang-algeb ra (YY 8 ) CAUG 312 for the genomatrix P CAUG 312 (3) . The elements f 0 and m 2 are the quasi-real units in this algebra. f 0 f 1 m 2 m 3 f 4 f 5 m 6 m 7 f 0 f 0 f 1 m 2 m 3 f 4 f 5 m 6 m 7 f 1 f 1 - f 0 m 3 - m 2 - f 5 f 4 - m 7 m 6 m 2 f 0 f 1 m 2 m 3 f 4 f 5 m6 m 7 m 3 f 1 - f 0 m 3 -m 2 - f 5 f 4 - m 7 m 6 f 4 f 4 f 5 m 6 m 7 f 0 f 1 m 2 m 3 f 5 f 5 - f 4 m 7 - m 6 - f 1 f 0 - m 3 m 2 m 6 f 4 f 5 m 6 m 7 f 0 f 1 m 2 m 3 m 7 f 5 - f 4 m 7 - m 6 - f 1 f 0 - m 3 m 2 Figure 10. The multiplicat ion table of the basic elem ents of the octave Yin-Yang-algeb ra (YY 8 ) CAUG 132 for the genomatrix P CAUG 132 (3) . The elements f 0 and m 2 are the quasi-real units in this algebra. f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 f 0 f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 m 1 f 0 m 1 f 2 m 3 f 4 m 5 f 6 m 7 f 2 f 2 m 3 f 0 m 1 f 6 m 7 f 4 m 5 m 3 f 2 m 3 f 0 m 1 f 6 m 7 f 4 m 5 m 5 f 4 m 5 - f 6 - m 7 - f 0 - m 1 f 2 m 3 m 5 f 4 m 5 - f 6 - m 7 - f 0 - m 1 f 2 m 3 f 6 f 6 m 7 - f 4 - m 5 - f 2 - m 3 f 0 m 1 m 7 f 6 m 7 - f 4 - m 5 - f 2 - m 3 f 0 m 1 Figure 11. The multiplicat ion table of the basic elem ents of the octave Yin-Yang-algeb ra (YY 8 ) CAUG 213 for the genomatrix P CAUG 213 (3) . The elements f 0 and m 1 are the quasi-real units in this algebra. f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 f 0 f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 f 1 f 1 f 0 f 3 f 2 m 5 m 4 m 7 m 6 f 2 f 2 - f 3 - f 0 f 1 m 6 - m 7 - m 4 m 5 f 3 f 3 - f 2 - f 1 f 0 m 7 - m 6 - m 5 m 4 m 4 f 0 f 1 f 2 f 3 m 4 m 5 m 6 m 7 m 5 f 1 f 0 f 3 f 2 m 5 m 4 m 7 m 6 m 6 f 2 - f 3 - f 0 f 1 m 6 - m 7 - m 4 m 5 m 7 f 3 - f 2 - f 1 f 0 m 7 - m 6 - m 5 m 4 Figure 12. The multiplicat ion table of the basic elem ents of the Yin-Yang-algebra (YY 8 ) CAUG 321 for the genomatrix P CAUG 321 (3) . The elements f 0 and m 4 are the quasi-real units in this algebra. Figures 9-12 dem onstrate the multiplications tabl e s of the basic elem ents of the Yin-Yang algebras for the other genomatrices P CAUG 312 (3) , P CAUG 132 (3) , P CAUG 213 (3) , P CAUG 321 (3) from the family of the six permutation genomat Taking into account the multiplication ta bles on Figures 6, 8-12 the proper YY 8 -numbers in the vector form of their presentation have the following expressions: (YY 8 ) CAUG 123 = x 0 * f 0 +x 1 * m 1 +x 2 * f 2 +x 3 * m 3 +x 4 * f 4 +x 5 * m 5 +x 6 * f 6 +x 7 * m 7 (YY 8 ) CAUG 231 = x 0 * f 0 +x 1 * f 1 +x 2 * f 2 +x 3 * f 2 +x 4 * m 4 +x 5 * m 5 +x 6 * m 6 +x 7 * m 7 (YY 8 ) CAUG 312 = x 0 * f 0 +x 1 * f 1 +x 2 * m 2 +x 3 * m 3 +x 4 * f 4 +x 5 * f 5 +x 6 * m 6 +x 7 * m 7 (YY 8 ) CAUG 132 = x 0 * f 0 +x 1 * f 1 +x 2 * m 2 +x 3 * m 3 +x 4 * f 4 +x 5 * f 5 +x 6 * m 6 +x 7 * m 7 (YY 8 ) CAUG 213 = x 0 * f 0 +x 1 * m 1 +x 2 * f 2 +x 3 * m 3 +x 4 * f 4 +x 5 * m 5 +x 6 * f 6 +x 7 * m 7 (YY 8 ) CAUG 321 = x 0 * f 0 +x 1 * f 1 +x 2 * f 2 +x 3 * f 3 +x 4 * m 4 +x 5 * m 5 +x 6 * m 6 +x 7 * m 7 (4) All these Yin-Yang matrices have secret conne ctions with Hadamard m atrices: when all their coordinates are equal to the re al unit 1 (x 0 =x 1 =…=x 7 =1) and when the change of signs of some components of the matrices takes place by means of the U-algorithm described in the article [Petoukhov, arXiv:0802.3366], then all these Yin-Yang octave matrices become the Hadamard matrices. In necessary cases biological computers of organism s can transform these Yin-Yang matrices into the Hadamard m atrices to operate with systems of orthogonal vectors. One can add that for the case when a ll their coordinates are equal to 1 (x 0 =x 1 =…=x 7 =1), all these six Yin-Yang matrices (YY 8 ) CAUG 123 , (YY 8 ) CAUG 231 , …, (YY 8 ) CAUG 321 possess the property of their tetra-reproduction which was descri which reminds the tetra-reproduction of gametal cells in the biological process of meiosis. One can mention two facts else. The comple mentary triplets (codon and anti-codon) play essential role in the genetic code system s. One can replace each codon in the genom atrices Р CAUG 123 , Р CAUG 231 , Р CAUG 312 , Р CAUG 132 , Р CAUG 213 , Р CAUG 321 by its anti-codon. The new six genomatrices appear in this case. Have they any connection with Yin-Yang algebras? We have investigated this question with the positive re sult. The multiplica tion tables for the basic elements of Yin-Yang matrices, connected with these new genomatrices, are iden tical to the multiplication tables for the initia l genomatrices. In other words, the “com plementary” transformations of the genomatrices Р CAUG 123 , Р CAUG 231 , Р CAUG 312 , Р CAUG 132 , Р CAUG 213 , Р CAUG 321 change the matrix forms of the initia l YY 8 -numbers only but do not change the Yin-Yang algebras of the genomatrices. But if we consider the transposed matrices, which are received from the matrices (YY 8 ) CAUG 123 , (YY 8 ) CAUG 231 , etc., they correspond to new Yin-Yang octave algebras. 4 The genetic Yin-Yang octaves as “double genoquaternions” We shall name any numbers with 8 item s x 0 * i 0 +x 1 * i 1 +…x 7 * i 7 by the name “octaves” independently of m ultiplication tabl es of their basic elem ents. We shall nam e numbers with 4 items x 0 * i 0 +x 1 * i 1 +x 2 * i 2 +x 3 * i 3 by the name “quaternions” independently of m ultiplication tables of their basic elements (quaternions by Ham ilton are the special case of quaternion s). Let us analyze the expression (3) of the genetic octave YY 8 together with its multiplication table (Figure 5). If all male coor dinates are equal to zero (m 1 =m 3 =m 5 =m 7 ), this genetic octave YY 8 becomes the genetic quaternion g f : g f = x 0 * f 0 +x 2 * f 2 +x 4 * f 4 +x 6 * f 6 (5) The proper multiplication table for this quatern ion is shown on Figure 13 (on the lef t side). This table is received from the m ultiplication table for the algebra YY 8 (Figure 5) by nullification (by excision) of the columns and rows , which have the male basic el ements. Taking into account that the basic element f 0 possesses the multiplication properties of the real unit relative to all female basic elements, one can rewrite the e xpression (5) in th e following f orm: g f = x 0 * 1 +x 2 * f 2 +x 4 * f 4 +x 6 * f 6 (6) If all female coordinates are equal to zero (f 0 =f 2 =f 4 =f 6 ), this genetic octave YY 8 becomes the genetic quaternion g m : g m = x 1 * m 1 + x 3 * m 3 + x 5 * m 5 + x 7 * m 7 (7) The appropriate multiplication table f or this quaternion is shown on Figure 13 (on the right side). Taking into acc ount that the basic element m 1 possesses the multiplication properties of the real unit relative to all m ale basic elem ents, one can rewrite the expressio n (7) in the following form: g m = x 1 * 1 + x 3 * m 3 + x 5 * m 5 + x 7 * m 7 (8) f 0 f 2 f 4 f 6 m 1 m 3 m 5 m 7 f 0 f 0 f 2 f 4 f 6 m 1 m 1 m 3 m 5 m 7 f 2 f 2 -f 0 -f 6 f 4 m 3 m 3 -m 1 -m 7 m 5 f 4 f 4 f 6 f 0 f 2 m 5 m 5 m 7 m1 m 3 f 6 f 6 - f 4 - f 2 f 0 m 7 m 7 - m 5 - m 3 m 1 Figure 13. The multiplication tables for the genetic quaternio ns g f (on the left side) and g m (on the right side). The quaternions g f and g m are similar to each other. They can be expressed in the following general form, the multiplica tion table of which is shown on Figure 14 (on the r ight side): g = y 0 * 1 + y 1 * i 1 + y 2 * i 2 + y 3 * i 3 (9) Figure 14 shows the comparison be tween the multiplication tables for q uaternions by Ham ilton (on the left side) and for these genetic qua ternions g (or briefly “genoquaternions”). 1 i 1 i 2 i 3 1i 1 i 2 i 3 1 1 i 1 i 2 i 3 11 i 1 i 2 i 3 i 1 i 1 -1 i 3 - i 2 i 1 i 1 -1 - i 3 i 2 i 2 i 2 - i 3 -1 i 1 i 2 i 2 i 3 1 i 1 i 3 i 3 i 2 -i 1 -1 i 3 i 3 - i 2 - i 1 1 Figure 14. The multiplication tables for quate rnions by Hamilton (on the left side) and for genoquaternions (on the right side). Quaternions by Hamilton q = x 0 *1 + x 1 *i 1 + x 2 *i 2 + x 3 *i 3 Genoquaternions g = x 0 *1 + x 1 *i 1 + x 2 *i 2 + x 3 *i 3 (q 1 *q 2 )*q3 = q 1 *(q 2 *q3) (g 1 *g 2 )*g3 = g 1 *(g 2 *g3) Conjugate quaternion q s = x 0 *1 - x 1 *i 1 - x 2 *i 2 - x 3 *i 3 Conjugate genoquaternion g s = x 0 *1 - x 1 *i 1 - x 2 *i 2 - x 3 *i 3 To the norm of quaternions: |q| 2 = q*q s = q s *q = x 0 2 + x 1 2 + x 2 2 + x 3 2 To the norm of genoquaternions: |g| 2 = g*g s = g s *g = x 0 2 + x 1 2 - x 2 2 - x 3 2 The inverse quaternion exists : q -1 = q s /|q| 2 The inverse genoqua ternion exists: g -1 = g s /|g| 2 (q 1 + q 2 ) s = (q 1 ) s + (q 2 ) s (g 1 + g 2 ) s = (g 1 ) s + (g 2 ) s (q 1 *q 2 ) s = (q 2 ) s * (q 1 ) s (g 1 *g 2 ) s = (g 2 ) s * (g 1 ) s Figure 15. The comparison of som e properties betw een the s ystems of quaternions by Hamilton (on the left side) and of genoqua ternions (on the right side). The system of quaternions by Hamilton has m a ny useful properties and applications in mathematics and physics. The author has receiv ed the essential result that the system of genoquaternions possesses many anal ogical properties, which permits to think about its useful applications in bioinformatics, mathem atical bi ology, etc. For example, the numeric system of genoquaternions is the system w ith the operation of division a nd it possesses the associative property, the notions of the “norm of genoquatern ion” and of the “invers e genoquaternion”, etc. Figure 15 demonstrates some analogies between both types of quaternions. Taking into account the expressions (5-9), one can name the genetic octave x 0 * i 0 +x 1 * i 1 +…x 7 * i 7 (with its individual multiplication table) as “the double gen oquaternion”. This name generates heuristic associations with the famo us name “the double spiral” of DNA. 5 The comparison between the classical ve ctor calculation and the genovector calculation The theory of quaternio ns by W.Hamilton possesses m any useful results and applications. Let us remind about one of them, which concerns the beautiful connectio n between these quaternions q = x 0 * 1 +x 1 * i 1 +x 2 * i 2 + x 3 * i 3 and the classical vector calcu lati on developed by J.Gibbs. One can take two vectors a and b , which belong to the plane ( i v , i w ), where v < w, v = 1, 2; w = 2, 3; a = a 1 *i v + a 2 * i w , b = b 1 * i 1 + b 2 * i 2 . These vectors can be presented in the following usual form: a = | a |*(i v *cos α + i w *sin α ), b = | b |*(i v *cos β + i w *sin β ), (10) where α and β are appropriate angles between these vectors and the axises i v and i w in the orthogonal system of the basic vectors ( i 1 , i 2 , i 3 ). If we multiply together thes e vectors as Hamilton’s quaternions in accord ance with the mu ltiplicatio n table on Figure 14 (the left side ), the following equation arises : a * b = - | a |*| b |*cos ( α – β ) + | a |*| b |*si n( α – β )* i vw , (11) where i vw is the third basic vector, which is orthogonal to the basic vectors i v and i m . The expression (11) shows that the quaternion mu ltiplica tion of two vectors conta ins two parts: the scalar part and the vect or part. The scalar p art | a |*| b |*c os( α – β ) is famous under the name “the scalar product” and the vector part a |*| b |*sin( α – β )* i 3 is famous under the name “the vector product” in the classical vector calculation. This vector calcula tion is utilized widely in mechanics to describe movements of hard bodies in our physical space, etc. Mechanics of bodies in the usual physical space fits this vector ca lculation. From the viewpoint of this vecto r calculation, the space is isotropic one becau se the e xpression (11) with its scalar and vector parts is the same for each of p airs of vectors, which belong to the p lanes ( i 1 , i 2 ), ( i 1 , i 3 ), ( i 2 , i 3 ), and the scalar products and the vectors product possess the analogical form s for all three cases of the planes. But what results arise, if we multiply togethe r the vectors a and b (10) as genoquaternions in accordance with their multiplication table (Figure 14, on the right s ide)? Let us consider the following three cases, each of which contains a s calar part and a vector part in the final expressions (12), (13), (14) , but in different forms. The case 1 . The vectors a and b belong to the plane ( i 1 , i 2 ). They can be expressed in the following form: a = | a |*( i 1 *cos α + i 2 *sin α ), b = | b |*( i 1 *cos β + i 2 *sin β ). If we multiply together these vectors as genoquaternions (Figur e 14, in the right side), the result arises: a * b = | a |*| b |*( i 1 *cos α + i 2 *sin α )*( i 1 *cos β + i 2 *sin β ) = - | a |*| b |*cos( α + β )+ | a |*| b |*s in( α - β )* i 3 (12) The expression (12) of the genovector calculati on differs from the expression (11) of the classical vector calculation in the scalar part only (by the value cos( α + β )). The case 2 . The vectors a and b belong to the plane ( i 1 , i 3 ): a = | a |*( i 1 *cos α + i 3 *sin α ), b = | b |*( i 1 *cos β + i 3 *sin β ). The product of these two vect ors as genoquaternions gives the following result: a * b = | a |*| b |*( i 1 *cos α + i 3 *sin α )*( i 1 *cos β + i 3 *sin β ) = -| a |*| b |*cos( α + β ) - | a |* | b |*sin( α - β )* i 2 (13) The expression (13) of the genov ector calculation differs from the classical expres sion (11) in the scalar part (by the value cos( α + β )) and in the vector pa rt (by the opposite sign). The case 3 . The vectors a and b belong to the plane ( i 2 , i 3 ): a = | a |*( i 2 *cos α + i 3 *sin α ), b = | b |*( i 2 *cos β + i 3 *sin β ). The product of these two vectors as genoquaternions gives a * b = | a |*| b |*( i 2 *cos α + i 3 *sin α )*( i 2 *cos β +i 3 *sin β ) =+| a |*| b |*cos( α - β ) - | a |*| b |*sin( α - β )* i 1 (14) The expression (14) of the genov ector calculation differs from th e classical expression (11) by the opposite sign in the scalar pa rt and in the vector part. We name vectors, which are considered as qen oquaternions (with applications of the rules of genoquaternion operations to them), as “genov ectors”. It is obvious that the genovector calculation fits the case of an an isotropic space because th e results of multiplication of arbitra ry vectors a and b depend on the plane, to which these vectors belong. T he spaces of biological phenomena of morphogenesis, growth, etc. ha ve anisotropic characters also . Since the genovector calculation was develope d by the author from the geneti c code features, it seems that this calculation (and its generaliz ation for the system of Yin-Ya ng genooctaves) can be adequate to model anisotropic processes in biological spaces including pro cesses of bioinform atics and of biological morphogenesis on different le vels of each united organism . Many mathematical form alisms and notions, which we re rec eived in the th eory of quaternions by Hamilton and which were utilized in many scientific branches, have their analogies in the theo ry of genoquaternions (and in the theory of genetic tetrions described be low). Details of these analogies and their possible app lications in differe nt scientific bran ches will be described in our next publications. 6 The parametric reduction of th e genetic octave Yin-Yang-algebra to the 4-dimensional algebra of tetrions. About the pre-code and its alphabet The previous paragraphs have c onsidered the numeric system YY 8 = x 0 * f 0 +x 1 * m 1 +x 2 * f 2 +x 3 * m 3 +x 4 * f 4 +x 5 * m 5 +x 6 * f 6 +x 7 * m 7 (3) with 8 arbitr ary coordinates x 0 , x 1 , …, x 7 . But in accordance with the matrix on Figure 3 all thes e 8 coordinates are exp ressed by means of 4 parameters α , β , γ , δ : x 0 = ααγ ; x 1 = ααδ ; x 2 = αβγ ; x 3 = αβδ ; x 4 = βαγ ; x 5 = βαδ ; x 6 = ββγ ; x 7 = ββδ (15) Hence these 8 coordinates are not independent on each other and they ar e interconnected by the following expressions: х 1 = х 0 * δ / γ ; х 3 = х 2 * δ / γ ; х 5 = х 4 * δ / γ ; х 7 = х 6 * δ / γ (16) One can see from the expression (15) that the c oordinates belong to the fe male (m ale) type if they have the symbol γ ( δ correspondingly) on their th ird position . The expressions (16) show the existence of the pairs of “complementary ” ma le and fem ale coordinates which differ by the coefficient δ / γ only: х 1 and х 0 ; х 3 and х 2 ; х 5 and х 4 ; х 7 and х 6 . These interconnections of coordinates lead to the particul ar form of the octave number YY 8 , where female coordinates х 0 , х 2 , х 4 , х 6 exist only (another possible form has the male coordinates х 1 , х 3 , х 5 , х 7 only): T = x 0 *( f 0 + δ / γ * m 1 ) + x 2 *( f 2 + δ / γ * m 3 ) + x 4 *( f 4 + δ / γ * m 5 ) + x 6 *( f 6 + δ / γ * m 7 ) = = ααγ * 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ + αβγ * 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 + βαγ * 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 0 0 1 δ / γ 0 0 0 0 0 0 1 δ / γ 0 0 0 0 + ββγ * 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 -1 - δ / γ 0 0 0 0 0 0 (17) Each of these four matrices ( f 0 + δ / γ * m 1 ), ( f 2 + δ / γ * m 3 ), ( f 4 + δ / γ * m 5 ), ( f 6 + δ / γ * m 7 ) is constructed by means of fusion of male and fem ale basic matric es of the complem entary pair into united object. It is interesting that the se four matrices form their own closed set relative to multiplic ation. Figure 16 shows the table of m ultiplication of these m atrices. f 0 + δ / γ * m 1 f 2 + δ / γ * m 3 f 4 + δ / γ * m 5 f 6 + δ / γ * m 7 f 0 + δ / γ * m 1 (1+ δ / γ )*( f 0 + δ / γ * m 1 ) (1+ δ / γ )*( f 2 + δ / γ * m 3 ) (1+ δ / γ )*( f 4 + δ / γ * m 5 ) (1+ δ / γ )*( f 6 + δ / γ * m 7 ) f 2 + δ / γ * m 3 (1+ δ / γ )*( f 2 + δ / γ * m 3 ) -(1+ δ / γ )*( f 0 + δ / γ * m 1 ) -(1+ δ / γ )*( f 6 + δ / γ * m 7 ) (1+ δ / γ )*( f 4 + δ / γ * m 5 ) f 4 + δ / γ * m 5 (1+ δ / γ )*( f 4 + δ / γ * m 5 ) (1+ δ / γ )*( f 6 + δ / γ * m 7 ) (1+ δ / γ )*( f 0 + δ / γ * m 1 ) (1+ δ / γ )*( f 2 + δ / γ * m 3 ) f 6 + δ / γ * m 7 (1+ δ / γ )*( f 6 + δ / γ * m 7 ) -(1+ δ / γ )*( f 4 + δ / γ * m 5 ) -(1+ δ / γ )*( f 2 + δ / γ * m 3 ) (1+ δ / γ )*( f 0 + δ / γ * m 1 ) Figure 16. The table of multiplic ation of the matrices ( f 0 + δ / γ * m 1 ), ( f 2 + δ / γ * m 3 ), ( f 4 + δ / γ * m 5 ), ( f 6 + δ / γ * m 7 ), which are basic elem ents of the genetic tetrions. In this reason the expression (17 ) with all possible values of real numbers α , β , γ , δ represents the new system of 4-dimensional numbers, which are nam ed “genetic tetrions ” (or genotetrions) to distinguish them from 4-dimensional hypercompl ex num bers called “quaternions” traditionally (including genoquaternions described above). If quaternions and other hypercom plex numbers have the real unit among their ba sic elem ents, tetrions have not the real unit among their basic elements at all. The first basic element ( f 0 + δ / γ *m 1 ) of the tetrions (17) is the m atrix presentation of the real number (1+ δ / γ ). This bas ic element p ossesses the commutative property r elative to all these basic elements. The first item x 0 *( f 0 + δ / γ * m 1 ) is considered as the scalar part of tetr ions, and other three items x 2 *( f 2 + δ / γ * m 3 ) + x 4 *( f 4 + δ / γ * m 5 ) + x 6 *( f 6 + δ / γ * m 7 ) are considered as the vector pa rt of tetr ions. The square of any basic element of the tetrions T is equal to (1+ δ / γ )*( f 0 + δ / γ * m 1 ) with the sign “+” or “-“. This peculiarity is demonstrated on Figure 16 in cells (marke d by bold borders) along the main diagonal. So instead of the real unit, tetrions have the real number (1+v), where “v” is the real number, which is equal to δ / γ in the case of the genetic tetrions T (17). One can consider such tetrions as the special generalization of appropriate hypercomplex num bers by means of utilizing any kind of real numbers in the role of their first basic elem ent instead of utilizing the real unit in this role in the case of traditiona l hypercom plex numbers. (Concerning to such form of generalization of hypercomplex numbers, for exam ple one can write the matrix p resentation of similar g eneralizatio n of com plex numbers: y 0 *[1, v, 0, 0; 1, v, 0, 0; 0, 0, 1, v; 0, 0, 1, v]+y 1 *[0, 0, 1, v; 0, 0, 1, v; -1, -v, 0, 0; -1, -v, 0, 0]). The system of tetrions T (17) possesses the co m mutative and associativ e properties. It is the system with operation of division from the left si de and from the right side (by analogy with the division in the system of quaternions). By definition the conjugate tetrion T S is presented by the expression: T S = x 0 *( f 0 + δ / γ * m 1 ) - x 2 *( f 2 + δ / γ * m 3 ) - x 4 *( f 4 + δ / γ * m 5 ) - x 6 *( f 6 + δ / γ * m 7 ) (18) The following expressions for two tetrions T 1 and T 2 hold true: (T 1 + T 2 ) S = (T 1 ) S + (T 2 ) S ; (T 1 *T 2 ) S = (T 2 ) S * (T 1 ) S (19) The square of the module of tetrions: |T| 2 = T*T S = T S *T = (1+ δ / γ )*(x 0 2 +x 2 2 –x 4 2 –x 6 2 ) = (1+ δ / γ )*[( ααγ ) 2 +( αβγ ) 2 –( βαγ ) 2 –( ββγ ) 2 ] (20) The inverse genotetrion exists: T -1 = T S /|T| 2 . It permits to define the operation of division traditionally by m eans of multiplication by th e inverse genotetrion. One can see that these properties of the genetic tetrion s are similar to the properties of geno quaternions considered above (Figure 15) and that the genotetrion’s an d genoquaternion’s tables of multiplication are similar to each other by the disposition of the signs “+” and “-“ (Figures 14 and 16). The system of genetic tetrions le ads to specia l kind of vector calculatio n. By analogy with the expressions (12-14) for genoquaternions, one can receive the similar expressions (21-23) of vector calculation for genotetrions. Let us analyse the m ultiplication of two vectors a and b (10) as tetrions in accordance with the multiplicatio n table (16) in the same three cases which were described in the section 5. In the result we receive the following expressions (21-23). The case 1 . The vectors a and b belong to the plane of the basic vectors ( f 2 + δ / γ * m 3 , f 4 + δ / γ * m 5 ). Then a * b = - | a |*| b |* (1+ δ / γ ) 2 *cos( α + β ) + | a |*| b |*sin( α - β ) *(1+ δ / γ ) * ( f 6 + δ / γ * m 7 ). (21) The case 2 . The vectors a and b belong to the plane ( f 2 + δ / γ * m 3 , f 6 + δ / γ * m 7 ). Then a * b = - | a |*| b |* (1+ δ / γ ) 2 *cos( α + β ) - | a |*| b |*sin( α - β ) *(1+ δ / γ )*( f 4 + δ / γ * m 5 ). (22) The case 3 . The vectors a and b belong to the plane ( f 2 + δ / γ * m 3 , f 6 + δ / γ * m 7 ). Then a * b = +| a |*| b |* (1+ δ / γ ) 2 *cos( α - β ) - | a |*| b |*sin( α - β )* (1+ δ / γ )*( f 2 + δ / γ * m 3 ). (23) It is obvious that the vector calc ulation of genetic tetrions fits the case of an anisotropic space because the results of multiplic ation of arbitrary vectors a and b depend on the plane, to which these vectors belong. Can the scalar and vector parts of genetic tetrions be considered correspondingly as the time coordinate and the sp ace coordinates in the theory of the genetic space-time? This and other interesting questions are under investigation now and their answers should be published later. In the described approach, the gene tic code is presented as the replic a of the tetrions in the ir matrix form. It permits to consider the algebra of genetic tetrions as a candidate for the role of the mathematical system of genetic preceding c ode (the “pre-code” or the more fundamental code) relative to th e genetic code. Really, from a traditional viewpoint, a c ode is an aggregate of symbols which corresponds to elem ents of information. In our alge braic case, the speech is about the matrix system, sym bols of which can be confr onted with triplets and with other elements of the genetic code. In other words, the genetic co de can be encode d itself by sym bols of elements of the tetrion num erical system. Such tetrion pre-code has its own pre-code alphabet, which consists of the four letters α , β , γ , δ in contrast to the usual geneti c alph abet A, C, G, U/T. This set of the letters α , β , γ , δ can be named the alphabet of genetic alg ebras or the algebraic alph abet of the genetic code as well. A revealing such te trion pre-code as new numeric system can help with sorting, ordering and deeper understanding of the genetic information. It can help also to develop new effective methods of processing and transfer of information in many applied problems. Mathematical features of such pre-code can explain evolutionary features of the genetic code. One should emphasize that not only the (8x8)-matrix YY 8 (Figures 2 and 3), but each of its (4x4)-quadrants and each of its (2x2)-subquadrants define its own special algebras, if we take into account the coordinates x 0 , x 1 , …, x 7 and the algebraic alphabet α , β , γ , δ . It means that the genetic code is an ensemble of special multidim ensional algebras from such matrix v iewpoint. Details of this statement will be pub lished by the author separately. 7 About genetic mechanics In the beginning of the XIX century the followi ng opinion existed: the world possesses the single real geometry (Euclidean geometry) and the si ngle arithm etic. But this opinion was neglected after the discovery of non-Euclidean geometries and of quaternions by Ham ilton. The science understood that different natural systems can po ssess their own individual geometries and their own individual algebras (see this th eme in the book [Kline, 1980]). The exam ple of Hamilton, who has wasted 10 years in his attempts to solve th e task of description of transformations of 3D space by means of 3-dimensional algebras without a success, is the very demonstrative one. This example says that if a scientist does not guess right what type of algebr as is adequate for the natural system, which is investig ated by him, he can waste m any years without result by analogy with Hamilton. One can add that geometrical a nd physical-geometrical properties of separate natural systems (including laws of conservations, theories of oscillations and waves, theories of potentials and fields, etc.) can depend on the type of algebras which are adequate for th em. The fact, that the genetic code ha s led us to the algebra of geneti c tetrions (which is the special case of the genetic octave Yin-Yang-algebra), testif ies in favor of the importance of this algebra for each united organism. It seems that m any difficulties of modern science to understand genetic and biological systems are determined by approach es to these system s from the viewpoint of non-adequate algebras, which were developed formerly for other systems at all. In particular, the classical vector calculation, which plays the role of the important tool in classical mechanics and which fit geometrical properties of our physical space, can be inappr opriate for important biological phenomena. We put forward the hypothesis that a very special mechanics of biogenetic system s exists, which is connected with the vector cal culation of genetic tetr ions and with their generalization in the form of Yin-Yang octaves. We name it “genetic mechanics” because o f its relation with th e genetic code. Modern biomechanics is the set of applications of classical m echanics to model properties of living substances. In our opinion, such traditional bi omechanics are not adequate to many biological phenomena and it will b e replaced in many aspects by the genetic mechanics in future. We think that living substa nce lives in its own biological space which has specific algebraic and geometric properties. The hypothesis about a non-Euclidean geometry of living nature exists long ago [Vernadsky, 1965] but without a concrete definition of the type of such geometry. And how one can construct such geometry if biological organ isms – bacteria , birds, fis hes, plants, etc. - differ from each other so significantly in their morphogenetic and m any other features? The discovery of the genetic code, the basic elements of which are ge neral for all b iological organism s, has permitted to hope that such geometric and algebraic tasks can be solved by mean s of investigation of genetic code structures. Some results of such investigation are presented in this article. It happens frequently, that mathematicians construct a new beautiful abstract m athematics and then they search for opportunities of its application in different ar eas of natural sc iences. On the contrary, in our case the phenomen ology of the genetic code has le d the author un expectedly to the new mathematics of tetrions and Yin-Yang-octa ves. And we investigate form al features of this mathematics on the second stage only. The genetic code is the result of a gigantic experiment of the nature. This molecular code bears the imprint of a great set of known and unknown laws of the nature. In this connection, algebraic features of genetic structures are very essential to guess right a perspectiv e direc tion of development of algebraic bases of mathematical natural sciences in future. In our opinion, the tetrion algebra, the Yin-Yang-algebra and their geometries can be useful not only for bi ology, but also for other fields of m athematical natural sciences and for applied sciences (signals processing, mathematical economy, etc.). For example, they permit to develop new algorithms and m ethods of dig ital signal processing. 8 The genetic code as the multidimensional num ber and the idea by Pythagoras The notion “number” is the main notion of m athem atics. In accordance with the famous thesis, the complexity of civilization is ref lected in complexity of numbers which are utilized by the civilization [Mathematics in the modern world, 1964]. "Number - one of the m ost fundamental concepts not only in mathem atics, but also in all natural scien ces. Perhaps, it is the more prim ary concept than such global categories, as time, space, substance or a field" [Hypercom plex numbers in geometry and in physics, 2004]. After establishment of real numbers in the hi story of developm ent of the notion “number”, complex numbers x 0 +i*x 1 have appeared. These 2-dimensional numbers have played the role of the magic tool for developm ent of theories and calculatio ns in the field of problems of heat, light, sounds, fluctuations, elasti city, gravitation, magnetism, elect ricity, current of liquids, the quantum-mechanical phenomena. It seems that m odern atomic stations, airplanes, rockets and many other things would not exist without knowledge of complex num bers because the appropriate physical theories are based on these numbers. C.Gauss, J.Argand and С . Wessel have demonstrated that a plan e with its properties fits 2-dim ensional complex numb ers. W.Hamilton has proved that the properties of our 3-dimensi onal physical space fit m athematical properties of the special quaternions. Ham ilton’s quaternions have played the sign if icant role in the his tory of mathematical natural sciences as well. For exam ple, the classical vector calcu lation is deduced from the theory of these quaternions. The author supposes that genetic te trions, genoquaternions and Yin-Yang octaves (“double genoquaternions”), which are prompted by the genetic code and which are new kind of generalized numbers, ca n play a significant role in appropriate sc ientific fields in future also. Pythagoras has formulated the famous idea: “A ll things are numbers”. This idea had a great influence on mankind. B.Russel wrote that he di d not know any other person who has influenced with such power on other people in the field of thought [Russel, 1945]. In this relation the world does not know the more fundamental scientific id ea than the idea by Pythagoras (it should be mentioned that the notion “num ber” was perfected in science after Pythag oras in the direction of generalized numbers such as hypercomplex numbers). W e reveal the fact that the genetic code with its degeneracy properties f its genetic te trions (which are the special case of Yin-Yang octaves or “double genoquaternions ”) by analogy with th e fact that the 3D physical space fits Hamilton’s quaternions. One can say in this relati on that the genetic code is the tetrion num ber (in a certain sense). Our result s give additional materials to the great idea by Pythagoras. “Number” is the m ain notion of mathematics. In the result of our inves tigations of the genetic code as the bases of bioinformatics, we fi nd ourselves in the field of fundamentals of mathematics and m athematical natu ral sciences unexpectedly. It has many m athematical and heuristic consequences. 9 About the tetrion algebra of the gene tic code as the algebra of operators This brief paragraph pays attenti on to one of the most perspectiv e directions of understanding of the possible biological meanings of the genetic tetri on algebra and of the Yin-Yang-algebras. It is known that mathematics has deals not only with algebras of numbers but with algebras of operators also (see historical remarks in the book [Kliene,1980, Chapter VIII]). G.Boole has published in 1854 year his brilliant work about investigations of laws of thinking. He has proposed the Boole’s algebra of logics (or logical ope rators). Boole tried to construct such operator algebra which would reflect basic prop erties of human thinking. The Boole’s algebra plays a great role in the modern scienc e because of its connection s with many scientific branches: mathematical logic, the problem of arti ficia l intelligence, com puter technologies, bases of theory of probability, etc. In our opinion, the described algebra of tetrions and Yin-Yang-algebra of the genetic code can be considered not only as the algebr as of the numeric systems but as the algebra of proper logical operators of genetic systems als o. This direction of thoughts can lead us to deeper understanding of logic of biological systems including an adva nced variant of the id ea by Boole (and by some other scientists) about development of algebraic th eory of laws of thinki ng. The author plans to publish a possible variant of such genetic algebras of logical operators later. One can add that biological organisms have f amous possibilities to u tilize the same structures in multi-purpo se destinations. The genetic algebras can be utili zed by biological organism s in different purposes also. 10 What is life from the viewpoin t of algebra? The problem of algebraiz ation of bioinformatics Taking into account the great meaning of the genetic code for biological organisms, the described discovery of algebraic properties of th e genetic code gives the basis for investig ation of biological organizations from the algebraic viewpoint. Modern al gebra is the wide branch of mathematics. It possesses m any own theorems, app lications of which to genetic systems can give new vision in the field of theoretical biology. It is essential also that alge bra plays the great role in the modern theory of information encoding and of signal processing. All these facts provoke the high interest to the question: wha t is life from the view point of algebra? This question exists now in parallel with the old question from the famous book by E.Schrodinger: what is life from the viewpoint of physics? One can add that attem pts are known in modern theoretical physics to reveal info rmation bases of physics; in these attem pts information principles are consid ered as the most fundamental. Here one can mention as well the known problem of geometrization of physics, that is the problem of creation and interpretati on of physical theories in a form of theories of invariants of groups of transformations. Such general approach to different physical theories was very fruitful. One can hope that the problem of algebraization of bioinform atics (and of biology, which is connected closely with bioinformatics), that is understanding bioinfor m ation phenomena from the viewpoint of algebras of the ge netic code, will be useful also. Appendix A: Some additional ma thematical peculiarities of the genetic octave Yin-Yang-algebra YY 8 A.1 Interrelations between hypercomplex numbers and Yin-Yang numbers As it was mentioned above, Yin-Yang numbers (YY-numbers) can be considered as the generalization of hypercomplex numbers. Each of 2 n-1 -dimensional hypercomplex numbers can be transformed into the 2 n -dimensional YY-number by a speci al algorithm . An inverse application of this algo rithm to a 2 n -dimensional YY-number gene rates the ap propriate 2 n-1 - dimensional hypercomplex num ber. Accordi ng to this algorithm, if we have a (2 n x2 n )-matrix, which represents a 2 n -dimensional hypercom plex number, we should replace each component of this matrix by the (2x2 )-matrix [ х к х к +1 ; х к х к +1 ]. By this algorithm we have the tetra- reproduction of matrix components which rem inds the tetra-reproduction of gametal cells in the result of meiosis. In such reason this al gorithm has the conditional name “the m eiosis algorithm”. For example, if we have the (2x2)-matrix of the presentation of complex numbers, this m eiosis algorithm transforms it into (4x4)-m atrix of the presentation of 4-dimensional “Yin-Yang- complex” num bers KK 4 , which fit the special multiplication table of the appropriate YY 4 -algebra (Figure 17). Really, according to this algorithm , each component x 0 and x 1 of the initial matrix is replaced by the (2x2)-matri x of the mentioned type: х 0 =[y 0 y 1 ; y 0 y 1 ], x 1 =[y 2 y 3 ; y 2 y 3 ]. In the result we have YY-complex numbers КК 4 = y 0 * f 0 +y 1 * m 1 +y 2 * f 2 +y 3 * m 3 , where f 0 and m 1 are the female and male quasi- real units; f 2 и m 3 are the female and male im aginary units with the properties ( f 2 ) 2 = - f 0 , ( m 3 ) 2 = - m 1 . f 0 m 1 f 2 m 3 y 0 y 1 -y 2 -y 3 f 0 f 0 m 1 f 2 m 3 x 0 -x 1 y 0 y 1 -y 2 -y 3 m 1 f 0 m 1 f 2 m 3 x 1 x 0 ; y 2 y 3 y 0 y 1 ; f 2 f 2 m 3 - f 0 -m 1 y 2 y 3 y 0 y 1 m 3 f 2 m 3 - f 0 -m 1 Figure 17. The matrix form s of presentation of complex numbe rs (on the left side) and YY-complex numbers (in the m iddle). On the right side: the multiplica tion table for basic elements of the YY-complex num bers. By inverse application of this algorithm, one can receive a 4-dim ensional hypercomplex number from the genetic YY-number YY 8 . The YY-matrix YY 8 (Figure 2) contains 4 kinds of (2x2)-sub- quadrants, each of which has one of pairs of coordinates: x 0 and x 1 ; x 2 and x 3 ; x 4 and x 5 ; x 6 and x 7 . One can replace each such sub-qua drant by a separate coordinate: [x 0 x 1 ; x 0 x 1 ] = у 0 ; [x 2 x 3 ; x 2 x 3 ] = у 1 ; [x 4 x 5 ; x 4 x 5 ] = у 2 ; [x 6 x 7 ; x 6 x 7 ] = у 3 . In the result the (4x4)-matrix Q is appeared, which represents the genoquaternion Q = y 0 * 1 + y 1 * i 1 + y 2 * i 2 + y 3 * i 3 , which was considered above and which has i 1 2 = -1, i 2 2 = i 3 2 = +1. Figure 18 shows the matrix Q and the multiplication table for this genoquaternion. Th e genoquaternion Q rem inds coquaternions (or split-quaternions, or para-quaternions, or hyperbolic quaternions), introduced by J.Cockle in 1849 year ( http://en.Qikipedia.o rg/Qiki/Coquaternion ), but their mu ltiplication tables have differences. We name the number Q “genoquaternion of the first type”. (A genoquaternion of the second type is produced by the special permutat ion of colum ns of the matrix Q, which is connected with the perm utation of positions in genetic duplets [Petoukhov, 2008b, p.203]). 1 i 1 i 2 i 3 y 0 -y 1 y 2 -y 3 1 1 i 1 i 2 i 3 Q = y 1 y 0 -y 3 -y 2 i 1 i 1 -1 - i 3 i 2 y 2 -y 3 y 0 -y 1 i 2 i 2 i 3 1 i 1 -y 3 -y 2 y 1 y 0 i 3 i 3 - i 2 - i 1 1 Figure 18. The matrix form of presentation of the hyperc omplex num ber Q (on the left side); its multiplication table (on the r ight side). A.2 About the sub-algebras YY 2 and YY 4 of the genetic Yin-Yang octave algebra YY 8 Two squares are marked out by bold lines in the left top corner of the multiplication table on Figure 5 for the case of the genomatrix Р CAUG 123 . The first of these square s with its si ze (2x2) is the multiplication tab le of the basic elements of the 2-dimensional Yin-Yang a lgebra YY 2 . Figure 19 shows two matrix forms of presen tation of appropriate Yin-Yang numbers YY 2 . One of these forms [z 0 z 1 ; z 0 z 1 ] coincides with the structure of each (2x2)-sub-quadrant of the genomatrices on Figures 1-3 in relation of the disposition of the YY-coordinates x 0 , x 1 ,…, x 7 and amino acids with stop-signals. It testifies that the algebra YY 2 participates in the structural organization of the genetic code. The second square with its size (4x4) on Figure 5 is the multiplication table of the 4-dim ensional Yin-Yang algebra YY 4 . The appropriate Yin-Yang numbers YY 4 possess the following vector form: YY 4 = z 0 * f 0 +z 1 * m 1 +z 2 * f 2 +z 3 * m 3 and these numbers coincide with the Yin-Yang generalization of complex numbers (Figure 17). 1 F 0 1 M 1 z 0 z 1 z 0 -z 1 1 F 0 1 F 0 1 M 1 z 0 z 1 ;- z 0 z 1 ;1 M 1 1 F 0 1 M 1 Figure 19. Two matrix form s of a presentation of the 2-dimensional numbers YY 2 (on the left side); the multiplic ation table of the ba sic elements of the Yin-Yang algebra YY 2 . A.3 About the geometrical in terpretation of the 2-dimens ional Yin-Yang numbers YY 2 It is known that complex numbers have been wi dely recognized only af ter finding of their geometrical interpretation on the g eometric p lan e of complex variables. This plane was named “Gauss-Argand plane” according to names of th e mathematicians who have in troduced such plane. Whether it is possible to offer a substant ial geometrical interpre ta tion of the 2-dim ensional Yin-Yang numbers YY 2 ? Yes, it is possible. For this pur pose we introduce the plane of Yin- Yang variables (or YY-plane). It is an ordinary plane with the Yin-Yang system of Cartesian coordinates. This Yin-Yang system (or YY-system ) has the coordinate axises f and m , which play a role of female and male axises. By analogy with the case of complex numbers, each 2-dimensional YY-number is denoted on this Y Y-p lane by the point or by the vector. A product XX*ZZ of two Yin-Yang vectors, where XX = x 0 * f 0 + x 1 * m 1 and ZZ = z 0 * f 0 + z 1 * m 1 , possesses a geometric sense on such plane. Really, the resu lt of non-commutative multiplication of such two YY-vectors is equal to the second v ector with the scale coefficient, which is equal to the sum of coordinates of the first vector (Figure 20, on the left side). Th e same first vector-factor at multiplication with all other vectors of the plane or of a geometric figure leads to th eir identical scaling (Figure 20, on the right side). XX * ZZ =(x 0 * f 0 +x 1 * m 1 )*(z 0 * f 0 +z 1 * m 1 )=(x o +x 1 )*(z 0 * f 0 +z 1 * m 1 ) ZZ * XX =(z 0 * f 0 +z 1 * m 1 )*(x 0 * f 0 + 1 * m 1 )= (z 0 +z 1 )*(x 0 * f 0 +x 1 * m 1 ) Figure 20. The non-commutative multip lication of two Yin-Yang vectors (on the left side). A scaling of a geometric figure on the Yin-Yang plane (on the right side). It associates with the known biological phenom enon of volumetric growth of the living bodies, observed at the most different lines and branch es of biological evolu tion. Biological bodies are capable to the mysterious volumetric growth, oc curring in the cooperative way in all volume of the body or of its growing part. It is one of sharp differences of liv ing bodies from crystals with their surface growth occurring due to a local addition of new porti ons of substance on the surface of the crystal. In this connecti on, the Yin-Yang geometry is one of candidates for the role of the geometry of biological volumet ric growth. Other details of th e Yin-Yang geometry will be published later. A.4 About algebras with many qua si-real units (polysex algebras) It should be mentioned that the names “b isex algebra”, “bisex geom etry”, “bisex numbers”, etc. can be utilized as the synonyms of the names “Yin-Yang algebra”, “Yin-Yang geom etry”, “Yin- Yang numbers”. In some cases the utilization of these names can be more comfortab le but it depends on situations . For example it is comfortable in th e question about algebras with many quasi-real units. Such algebras can be named “polysex algebras” (or “n-sex algebras”). One can denote that bisex algeb ras are a partic ular case of n-sex algebras, each of which possesses a set of their basic elements with “n” qua si-rea l units but with out the real unit. Figure 21 shows the simplest example of 3-sex num bers x 0 * i 0 +x 1 * i 1 +x 2 * i 2 (in the matrix form of their presentation), which contain three quasi- real numbers only. T he basic elements i 0 , i 1 , i 2 of these 3-sex numbers have their matrix form s: i 0 =[1 0 0; 1 0 0; 1 0 0], i 1 =[0 1 0; 0 1 0; 0 1 0], i 2 =[0 0 1; 0 0 1; 0 0 1]. Their multiplica tion table is shown on Figure 21. i 0 i 1 i 2 x 0 x 1 x 2 i 0 i 0 i 1 i 2 x 0 x 1 x 2 ; i 1 i 0 i 1 i 2 x 0 x 1 x 2 i 2 i 0 i 1 i 2 Figure 21. The matrix form of presentation of 3-sex numbers and the multiplication table o f their basic elements. Bisex numbers and trisex numbers can be cons idered as numeric analogies of the fam ous symbols Yin-Yang and tomoe (Figure 22). Details a bout the Japanese tom oe symbol are given at the site http://altreligion.about.com /library/glossary/symbols/bldefstomoe.htm . ☯ Figure 22. The symbol Yin-Yang and the symbol tomoe. Multiplication of two 3-sexes gives the resu lt which is similar to the describ ed case of bisexes: the result is the 3-sex, which is equal to the seco nd factor increased by th e sum of coordinates of the first factor (Figure 23). The geometry of 3- sexes is a candidate to play the role of the geometry of the volumetric biological growth in the case of 3D-space (by analogy with the geometry of bisexes in the case of a plane). x 0 , x 1 , x 3 x 0 , x 1 , x 2 x 0 , x 1 , x 2 * y 0 , y 1 , y 3 y 0 , y 1 , y 2 y 0 , y 1 , y 2 = (x 0 +x 1 +x 2 )* y 0 , y 1 , y 3 y 0 , y 1 , y 2 y 0 , y 1 , y 2 Figure 23. Multiplication of two 3-sexes. Figure 24 shows another example of polysexes: the m atrix form of 8-dim ensional 4-sexes x 0 * i 0 +x 1 * i 1 +x 2 * i 2 +x 3 * i 3 +x 4 * i 4 +x 5 * i 5 +x 6 * i 6 +x 7 * i 7 , which have 4 quasi-real units i 0 , i 1 , i 2 , i 3 and which have its own imaginary unit for each of these quasi-real units: i 4 2 = - i 0 ; i 5 2 = - i 1 ; i 6 2 = - i 2 ; i 7 2 = - i 3 . x 0 x 1 x 2 x 3 –x 4 –x 5 –x 6 –x 7 x 0 x 1 x 2 x 3 –x 4 –x 5 –x 6 –x 7 x 0 x 1 x 2 x 3 –x 4 –x 5 –x 6 –x 7 x 0 x 1 x 2 x 3 –x 4 –x 5 –x 6 –x 7 x 4 x 5 x 6 x 7 x 0 x 1 x 2 x 3 x 4 x 5 x 6 x 7 x 0 x 1 x 2 x 3 x 4 x 5 x 6 x 7 x 0 x 1 x 2 x 3 x 4 x 5 x 6 x 7 x 0 x 1 x 2 x 3 i 0 i 1 i 2 i 3 i 4 i 5 i 6 i 7 i 0 i 0 i 1 i 2 i 3 i 4 i 5 i 6 i 7 i 1 i 0 i 1 i 2 i 3 i 4 i 5 i 6 i 7 i 2 i 0 i 1 i 2 i 3 i 4 i 5 i 6 i 7 i 3 i 0 i 1 i 2 i 3 i 4 i 5 i 6 i 7 i 4 i 4 i 5 i 6 i 7 -i 0 -i 1 -i 2 -i 3 i 5 i 4 i 5 i 6 i 7 -i 0 -i 1 -i 2 -i 3 i 6 i 4 i 5 i 6 i 7 -i 0 -i 1 -i 2 -i 3 i 7 i 4 i 5 i 6 i 7 -i 0 -i 1 -i 2 -i 3 Figure 24. The matrix form of presentation of 8-dimensional 4-sexes (the upper matrix) and their multiplication table (the lower table) Descriptions of other mathemati cal properties of Yin-Yang algebras and their in terrelations with the bioinformation systems will be continued in our next publications . YY-numbers permit to develop new class of mathem atical models of se lf-reproduction systems. The author investigates bisex generalizations of known physical equati ons additionally to find new results with a physical sense from there (it is the mathemati cal fact that known physi cal equations can be received from appropriate bisex equations by passa ge to the lim it in values of appropriate bisex coordinates). The algebraic theory of the ge netic code can say many useful and unexpected things about an origin of the genetic code and about la ws of living substances. Appendix B: About the notion “algebra” The notion “algebra” has two main senses. Accordi ng to the f irst sense, which is famous m ore widely, the algebra is the whole section of ma them atics about mathematical operations with mathematical sym bols. According to the second sense, whic h is utilized in th is article, algebra is a mathematical ob ject with certain properties or , better to say, arithm etic of multidim ensional numbers. By definition in the frame of this se cond sense, algebra A with its dimension “n” over a field P is a set of expressions x 0 * i 0 +x 1 * i 1 +x 2 * i 2 +…+x n-1 * i n-1 (where x 0 , x 1 ,…, x n-1 belong P; i 0 , i 1 , …i n-1 are basic elements of vectors, which fit such express i ons). This set is provided with the operation of multiplication by elements “k” f rom the field P to determ ine the formula k*(x 0 * i 0 + x 1 * i 1 +x 2 * i 2 +…+ x n-1 * i n-1 ) = k*x 0 * i 0 + k*x 1 * i 1 + k*x 2 * i 2 +…+ k*x n-1 * i n-1 . This set is provide d with the following operation of addition also: (x 0 * i 0 +x 1 * i 1 +x 2 * i 2 +…+x n-1 * i n-1 ) + (y 0 * i 0 +y 1 * i 1 +y 2 * i 2 + …+y n-1 * i n-1 ) = (x 0 +y 0 )* i 0 + (x 1 +y 1 )* i 1 + … + (x n-1 +y n-1 )* i n-1 . This set is provided with the operation of multiplication between symbols i r else, which is given by a specific multiplication table i r *i v = w rv,0 *i 0 + w rv,1 *i 1 +… w rv,n-1 *i 2 . This multiplication table is utilized to find the result of multiplications (x 0 * i 0 +x 1 * i 1 +x 2 * i 2 +…+x n-1 * i n-1 )*(y 0 * i 0 +y 1 * i 1 +y 2 * i 2 + …+y n-1 * i n-1 ). Any algebra is defined com pletely by its multiplicatio n tab le that is by a certain set of numbers w rv,u . These numbers do not subordinate to any conditi ons, and any such set of numbers defines the certain algebra. Acknowledgments : Described researches were made by th e author in the frame of a long-term cooperation between Russian and Hungarian Acad emies of Sciences and in the fram e of programs of “International Society of Symmetry in Bioinformatics” (USA, http://polaris.nova.edu/MST/ISSB ) and of “International Symm etry Association” (Hungary, http://symmetry.hu/ ) . The author is grateful to Frolov K.V., Darvas G., Ne’eman Y., He M., Bakhtiarov K.I., Kassandrov V.V., Smolianinov V. V., Vladimirov Y.S. for their support. References 1. Chernavskiy, D.S. (2000) The problem of origin of life and thought from the viewpoint of the modern physics. – Uspehi phizicheskih nauk , v. 170, № 2, p. 157-183 (in Russian). 2. Dawkins, R . (1991) The blind Watchmake r, p.112 (originally published by Longman Scientific & T echnical, 1986) 3. He M., Petoukhov S.V. (2007) Harmony of living nature, symmetries of genetic systems and matrix genetics. International journal of integrative medicine , 1 , 1, 41-43. 4. Hypercomplex numbers in geometry and in physics , 1, 2004, p. 4, leading article (in Russian) 5. Kadison R.V ., Ringrose J.R . (1983) Fundamentals of the theory of operator algebras (vol. 1) Elementary Theory . New York: Academic Press. 6. Kline M . (1980). Mathematics. The loss of certainty. N-Y, Oxford University Press. 7. Konopelchenko B.G., Rumer Yu.B . (1975). Classification of the codons in the genetic code. DAN SSSR , 223 , #2, p.145-153 (in Russian). 8. Mathematical encyclopedia (1977), 1-5 volumes. Moscow , “Sovetskaya encyclopedia” (in Russian). 9. Mathematics in the modern world (1964). Scientific Am erican, N.-Y. 10. Petoukhov S.V. (2001a) Genetic codes I: binary s ub-alphabets, bi-symmetric m atrices and the golden section; Genetic codes II : numeric rules of degeneracy and the chronocyclic theory. Symmetry: Culture and Science , 12 , #3-4, p. 255-306. 11. Petoukhov S.V. (2003-2004) Attributive conception of genetic code, its bi-periodic tables and problem of unificati on bases of biological languages. Symmetry: Culture and Science , 14 - 15 , part 1, 281-307. 12. Petoukhov S.V. (2005) The rules of degeneracy and segregations in genetic codes. The chronocyclic conception and paralle ls with Mendel’s laws. - In: Advances in Bioinformatics and its Applications, Series in Math ematical Biology and Medicine, 8 , p. 512-532, World Scientific. 13. Petoukhov S.V. (2006a) Bioinformatics: m atrix geneti cs, algebras of the genetic code and bi о logical harmony. Symmetry: Culture and Science , 17 , #1-4, p. 251-290. 14. Petoukhov S.V. (2008a) The degeneracy of the gene tic code and Hadamard m atrices. arXiv:0802.3366 [q-bio.QM]. 15. Petoukhov S.V. (2008b) Matrix genetics, algebras of the genetic code, noise imm unity. M., RCD, 316 p. (in Russian) 16. Petoukhov S.V. (2008c) Matrix genetics, part 1: perm utations of positions in triplets and symmetries of genetic matrices. ar Xiv:0803.0888 [q-bio.QM q-bio.GN q-bio.OT] 17. Russel B. (1945) A History of Western Philosophy . N.Y. 18. Vernadsky V.I. (1965) Chemical structure of the Ea rth and its surrounding. Moscow , Nauka.
Original Paper
Loading high-quality paper...
Comments & Academic Discussion
Loading comments...
Leave a Comment