Virus Structure: From Crick and Watson to a New Conjecture
We conjecture that certain patterns (scars), theoretically and numerically predicted to be formed by electrons arranged on a sphere to minimize the repulsive Coulomb potential (the Thomson problem) and experimentally found in spherical crystals forme…
Authors: Alfredo Iorio, Siddhartha Sen
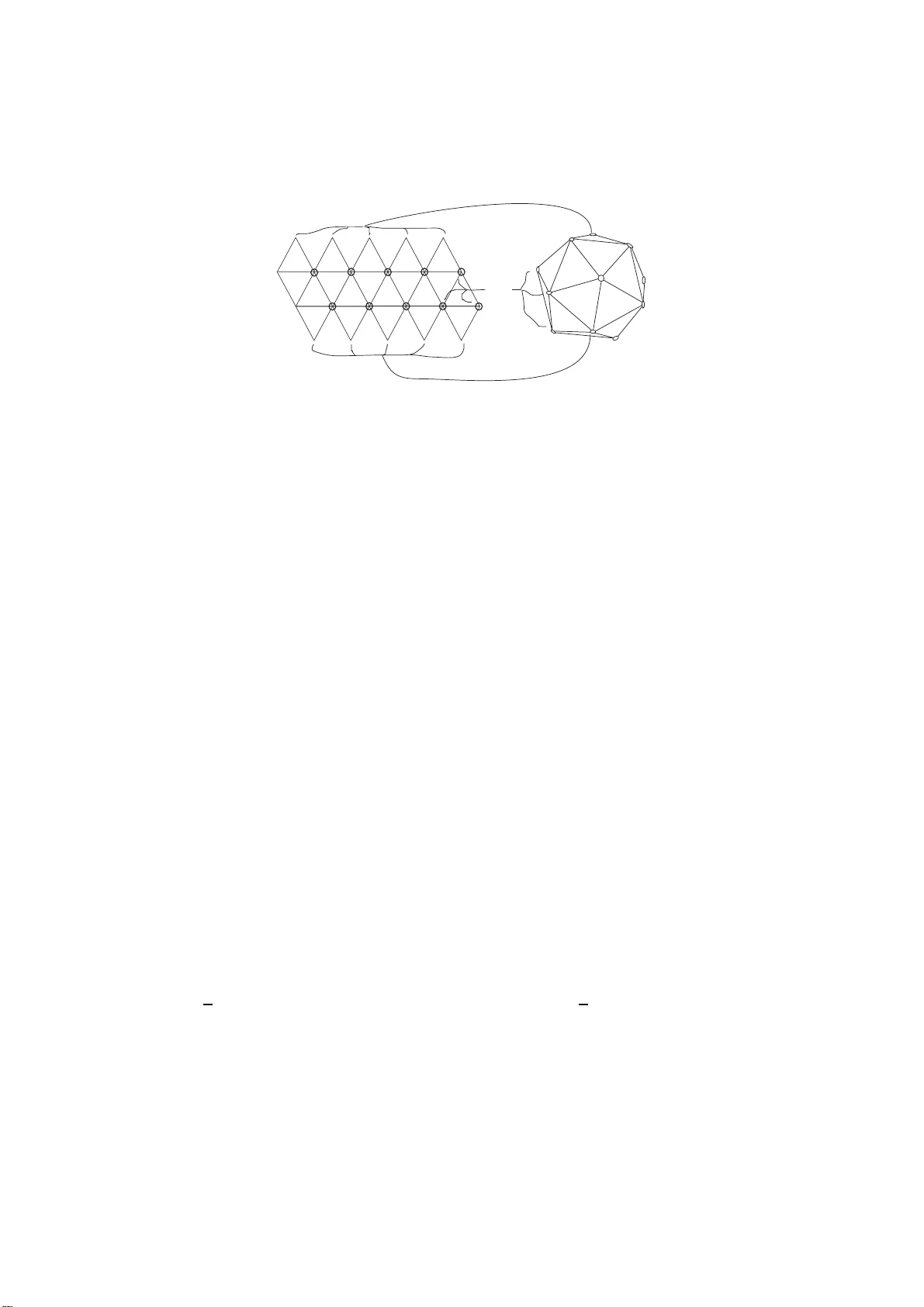
Vir us Str ucture: Fr om Crick and W a tson to a new conjecture ∗ Alfredo Iorio a,b 1 and Siddhar tha Sen c,d 2 a Institute of P article and Nuclear Ph ysics, Charles Univ ersit y of Prague V Holeso vic k ach 2, 18200 Prague 8 - Czec h Repub lic b Departmen t of Ph ysics “E.R.Caianiell o” Univ ersit y of Salerno and INFN Via Allende, 8408 1 Baronissi (SA) - Italy c Sc ho ol of Mathematical Science, Universit y College Dub lin Belfield, Dublin 4 - Ireland d Indian Asso ciation for the Cultiv atio n of Science Jada vpur, Calcutta 700032 - I ndia Abstract W e c onjecture that certa in patterns (sca r s), theoretically and n u- merically predicted to be formed by electro ns a rranged on a sphere to minimize the repulsive Coulomb potential (the Thomson pro blem) and exp erimentally found in spherical crystals formed by self-assembled po lystyrene beads (an instance of the gener ali ze d Thomson pro blem), could be relev ant to extend the clas s ic Caspar and Klug construction for icosa hedrally-shap ed virus ca psids. The main idea is that sca r s could b e produced at an intermediate stag e of the as semb ly of the virus ca psids and the rele a se of the b ending energy pre s en t in scar s int o stretching energy co uld allow for a v ariety of non-spher ical capsids’ shap es. The conjecture can be tested in exp eriments on the ass em bly of artificial protein-cages where these scars should app ear. ∗ In vited talk b y A.I. a t the F ourth In ternational Summer Sc ho ol and W o rk- shop on Nuclear Ph ysics Methods and Accelerators in Biology and Medicine - Prague, 8-19 July 2007. P A CS: 87.10.+e, 87.15.Kg, 61.72.- y Keyw ords: Viru s s tr ucture, Biomembranes, Cr ystal Defects 1 E-mail: iorio@ipnp.tro ja.mff.cuni.cz 2 E-mail: tcss@mahendra.iacs.res.in 1 Virus Structure General Considerations Viruses are small p ieces of genetic material (DNA or RNA) that can efficien tly encod e few small identi cal proteins that then assem ble themselv es 3 to f orm “cages” around the genetic material [1]. These cages are called c apsids and their shap e is the main concern of this pap er. Capsids are essentia l for protecting the genetic material and con tribute to identifying cells suitable for the duplication of the genetic material 4 . Un- derstanding the w a y p roteins arrange to form these v ery r esistent capsids is imp ortan t: i f we could find a wa y to u ndo these constructions we wo uld b e able to destro y viruses. There are th ree classes of capsid’s shap es [1]: he- lic al (the proteins sp iralize counter-c lo c kwise aroun d the genetic material) , ic osa he dr al or simple (the proteins arrange in mo rp h ologica l units of 5 and 6 follo wing pr ecise geometrical and top ological pr escriptions, as w e shall so on explain), c omplex (sphero-cylindrical, conical, tubu lar or eve n more compli- cated s h ap es, i.e. without a p recise resemblance to an y particular regular p olyhedron). There are also p oly morphic viruses th at change their shap e, e.g., from icosahedral to tubu lar and envelop e d viruses that, in addition to the protei n-capsid, also hav e an o uter lipid bila yer ( the viral en velope) tak en b y the host cell membranes. Icosahedral Viruses In 1956 Cr ick and W atson [2] prop osed that small viruses ha ve capsid s with the identica l proteins (or subunits or structur al units ) arranged in to morphological un its called c apsom ers with the shap e of hexagons and p ent agons, calle d hexamers and p entamers , resp ectiv ely . These capsomers form p olyhedrons that go under the name of i c osadelta- he dr ons , with a fixed n umber , 12, of p enta mers and a v ariable n umber of hexamers. F ollo win g Cric k a nd W a tson’s seminal idea, Caspar and Klug (CK) [3] later extended the class of viru ses to which this construction ap- plies to wh at they called “simple viru ses”, i.e. still rou gh ly spherical bu t not necessarily small viruses. The CK mo del f or icosadeltahedral capsids is no w ada ys un iv ersally accepted by virologists [1], [4]. The fact that exactly 12 p enta mers are n ecessary is easily und ersto o d if w e lo ok at this pr oblem as the analogue problem of tiling a sphere with p ent agons and hexagons and we tak e int o accoun t the top ological prop er- ties of the sphere (Euler theorem, see, e.g., [5]). The precise n umb er of hexamers will n ot b e fixed by this argument and needs further assumptions that we shall d iscuss in the next paragraph . Th e argument goes lik e th is: 3 Sometimes, for large viruses, the assem bly is done with the help of other proteins encod ed to this end by the genetic material. The environment also pla ys an important role. 4 Viruses are not able to duplicate without the help of the host cell, that is why their living nature is debatable. 1 a b k=3 h=1 n A Figure 1: Planar hexagonal lattice of id en tical rigid proteins. The ve ctor ~ A = h ~ a + k ~ b corresp onds to h = 1 and k = 3. Supp ose that th e ends of p r oteins only join three at the time. I f N p is th e n umb er of p -mers used to tile a sphere of unit radius, i.e. N 5 p ent amers and N 6 hexamers, the resu lting p olyhed ron P has V P = 1 / 3 P N N p p v er- tices, E P = 1 / 2 P N N p p edges, and F P = P N N p faces, giving for the Euler c haracteristic χ = V P − E P + F P , the follo wing expr ession X N (6 − p ) N p = 6 χ = 12 , (1) since for a sph ere χ = 2. Explicitly Eq.(1) reads (6 − 5) N 5 + (6 − 6) N 6 = 12 (2) hence to tile a sph ere N 5 = 12 is required , but N 6 can b e arb itrary . As said, for virus capsids N 6 is n ot arbitrary b ut m ust b e a sp ecific num b er that we shall so on obtain. F or the mathematical p r oblem of th e tiling of the sphere one m ight also imagine to u se heptagons . In that case the Euler formula (1) giv es N 5 − N 7 = 12 . (3) Th us, starting from the tiling of the sph ere with exactly 12 p entag ons (and an arb itrary n umber of hexagons) one can add p airs p en tagon-heptagon, but not a p en tagon or a heptagon separately . Note that at this p oin t this is only a mathematica l consideration and its relev ance for virus structure is all to b e pro v ed. The geometric interpretation of the Eu ler formula (1) is that a sphere of u nit r adius has curv ature R sphere = + 1 and eac h p olygon cont ribu tes to this curv ature with R p = (6 − p ) / 12: a hexagon with R 6 = 0, a p en tagon with R 5 = + 1 / 12, a heptagon with R 7 = − 1 / 12. This can b e und ersto o d by constructing hexagons, p entag ons and heptagons out of equilateral triangles of pap er. A hexagon is obtained by joining together 6 triangles: they all sta y in a p lane. T ak e one triangle ou t and join wh at is left to make a p entag on and the resulting figure will b en d outw ards, while adding one triangle to 2 1vertex (northpole) 1vertex (southpole) Inner vertices A A A A A A Figure 2: Th e equilateral triangles template and the icosadelta hedron. The 10 circled p oints on the planar template corresp ond to the 10 inner vertice s of the solid, w hile all the outer vertices of the 5 upp er triangles corresp ond to the n orth p ole vertex of the solid and all the outer ve rtices of the 5 lo w er triangles corresp ond to the south p ole v ertex. At these lo cations th e hexamers turn into p en tamers. Eac h triangular face of the icosadeltahedron is made of [ T / 2] hexamers (6 f or the example of Fig.1). the hexagon to mak e a hep tagon results in to an in w ard b ending. This also tells u s that a certain amoun t of b ending ener gy E b is necessary to con ve rt a hexagon into a p en tagon or in to a hep tagon. Ho w big is E b dep ends on the elastic prop erties of the material used. Let us no w describ e in more detail the CK construction. The CK construction Supp ose that the proteins are arranged on a plane to form th e h exagonal lattice of Fig.1. Eac h sid e of the lattice rep r esen ts a real p rotein. The basic v ectors ~ a and ~ b , with | ~ a | = | ~ b | , join the cen ter of the hexagon tak en as the origin of the lattice with the cen ters of the nearest hexagons as in figure. The angle is evid ently ϕ = 60 o . The 3-dimensional p olyhedron these proteins will ev entuall y f orm is obtained by imagining the 20 equilateral triangles with side | ~ A | = A - where ~ A = h ~ a + k ~ b , and h, k = 0 , 1 , 2 , ... - represen ted in Fig.2 folded to ob tain the icosahedron, the platonic solid with 12 vertic es, 20 faces and 30 edges. Eac h triangle face of the icosahedron, conta ins a fixed n umber of h examers that are the real proteins. A t eac h of th e 12 v ertices the h examers must tur n into p ent amers for the top ologic al and geometrical reasons describ ed ab ov e. Sa y | ~ a | = a , then one h as A 2 = a 2 ( h 2 + k 2 + 2 hk cos ϕ ) = a 2 ( h 2 + k 2 + hk ) ≡ a 2 T ( h , k ), with T ( h, k ) = 1 , 3 , 4 , 7 , ... . Being th e area of th e triangle giv en by α A = ( √ 3 / 4) a 2 T ( h , k ) and the area of one h exagon α 6 = ( √ 3 / 2) a 2 , the num b er of hexagons p er triangle is n 6 = α A /α 6 = [ T / 2]. T he total n umb er of sub units is obtained by counting the total num b er of hexagons used for the planar lattice of Fig.1, whic h is N 6 = 20( T / 2) = 10 T , then m ultiplying by 6 (the n umb er of edges of the hexagon): N proteins = 60 T . On the real 3-dimensional 3 N Proteins T F eline P anleuko p enia Virus 60 1 Human Hepatitis B 240 4 Infectious Bur sal Disease Viru s (IBD V) 780 13 Gener al 60 T h 2 + k 2 + hk T able 1: Examples of viruses th at follo w the CK classification tak en from Ref. [4]. solid (that one one m igh t th in k of obtaining by f oldin g the planar template) the 60 T p r oteins are arranged as: i) 60 form 12 p ent amers; ii) 60( T − 1) form 10( T − 1) hexamers, for a total n umb er of morp hologica l units of N = 10 T +2. The figures obtained are icosadeltahedrons c haracterized by the pair of intege rs ( h, k ) whic h n ot only are related to th e total num b er of pr oteins, but also giv e th e “chiralit y” of the p olyhedron. Viru ses b elonging to this class follo w these prescriptions with great accuracy and they are classified according to the v alues of T (see T able 1 for some examples and Ref.[4] for an exhaustiv e database on icosahedral virus structures). Recen tly there ha v e b een v arious attempts to generalize the CK mo del to include also certain complex viruses. One of those attempts is the mo del prop osed in Ref.[6] - based on the con tin uum elastic theory of large sp herical vir uses of Ref.[7] - wh ere the authors address the prob lem of u nderstanding the form ation of sp hero cylindrical and conical virus capsids. Later w e shall show that, if a c hange in the texture of th e arrangemen t of proteins (scar) tak es place, those and m any more shap es could b e obtained. 2 Lessons from the Th omson Problem Thomson Problem Let us no w tur n our atten tion to a differen t but ge- ometrically related physical set-up from whic h we would lik e to gain some insigh ts for the generalizat ion of the CK construction we are lo oking for: the Thomson p r oblem [8]. It consists of determinin g the minim um energy configuration for a collection of electrons constrained to mo v e on the sur face of a sph ere and int eracting via the Coulom b p otent ial. T his old (and largely unsolv ed) p roblem has man y generalizations for more general repulsive p o- ten tials as w ell as for top ological defects rather than u nit electric charge s [9], [10]. The fact that the t w o problems (virus capsid s construction and equilibrium configurations for c harges on a sphere) are intimat ely related can b e seen from the numerical resu lts for the Thomson pr oblem that ha v e b een obtained o ve r the y ears. In Ref. [11] the authors prop osed as solution of the problem an arrangement of N electrons on the sphere into a triangular lattice where eac h electron has 6 nearest neigh b ors sitting at the vertic es of an hexagon, with the exception of 12 lo cations where the nearest n eigh b ors 4 are only 5 sitting at the vertices of a p en tagon and N = 10 T + 2, with T = h 2 + k 2 + hk : th at is the icosadelta hedr on. Note that in this case the electrons are constrained to b e on the surface of the sphere, e.g. imagining the s phere as a metal, while the proteins h a v e not suc h constraint . F urther- more, th e p olygons h ere are “imaginary”, in the sense that only the vertic es are real p articles, while the edges are n ot. Scars (and P en tagonal Buttons) F urther stud ies [12] hav e shown that, ev en for N = 10 T + 2 electrons, when T is large enough (of the order of 10 2 ), configurations which differ from th e icosadeltahedron hav e lo wer en- ergy than th e corresp ond ing icosadeltahedron. That is, when near on e of the 12 p en tagons t w o h exagons (let u s call th is a 5-6-6 str u cture) are replaced by a pair heptagon-p en tagon (let us call this a 5-7-5 structur e) to f orm a linear pattern called sc ar , the energy is lo w er than th at of a configur ation of 12 p ent agons and all the rest hexagons. Th is ind eed happ ens in numerica l sim- ulations for higher and higher num b er of electrons, w h ere the s cars b ecome longer (e.g. 5-7-5-7-5, etc.) , alw a ys resp ect the top ological/geo metrical con- strain t of Eq. (3), can sp iralize or might ev en form exotic patterns lik e t w o nested p ent agonal structur es with fiv e p enta gons placed at the vertice s of the outer p entag onal stru cture, five heptagons at the v ertices of the inner p en - tagonal structure, and a p entag on in th e common cen ter (the vertex of the icosadeltahedron) (see, e.g., [9] and referen ces th er ein). The latter patterns are called p entagonal buttons and an exp lanation of th eir top ological origin can b e foun d in Ref.[5]. Ap p aren tly , ev en more complicated structures can app ear in n umerical sim ulations [9]. Scars ha v e b een exp erimen tally found to b e f ormed in sp herical cry s tals of mutually rep elling p olyst yrene b eads self-assem bled on wa ter droplets in oil [13]. The r epulsiv e p oten tial there is not the Coulomb p oten tial, h ence that is a particular in stance of the gener- alized Th omson pr oblem. Th ese exp erimen tal fin dings confirm that, at least in the case of scars, thin gs go along the lines of the ab o v e outlined analysis. The lesson we learn from the Th omson pr oblem is that und er certain conditions it is energeticall y fa v orable to con v ert a pair 6-6, with zero total and lo cal cur v ature (0 = 0 + 0) and zero b end ing energy , into a pair 5- 7, again w ith zero total curv ature b ut with nonzero lo cal curv ature (0 = +1 / 12 − 1 / 1 2) hence with nonzero b en ding ener gy giv en b y 2 E b , where E b is n ecessary to con ve rt a 6 into a 5 or into a 7. 3 Scars and Virus Structure Our Conjecture W hat we pr op ose here is that, du e to the in teraction with the environmen t (and/or with the genetic material) , the f ormation of scars of p enta mers and heptamers can tak e p lace in viru s capsids dur ing the pro cess of assem b ly of the proteins. The wa y we b eliev e this happ ens is as 5 follo w s: i) At fi rst th e p roteins assem ble to make an icosadeltahedron follo w- ing the CK prescrip tion. ii) A t an in termediate stage, d ue to the inte raction with the environmen t they form scars n ear the lo cation of one or m ore of the 12 p en tamers at the vertice s of th e icosadeltahedron. This interac tion is necessary b ecause the needed extra b ending energy (2 E b in the case of the formation of what we might call a “simple” scar: 5-7-5 ) can only come fr om the en vironment . iii) Eve ntuall y , the capsids change s h ap e, from spherical to non-spherical via the release of the b end ing energy int o stretc hing en- ergy at th e location of the scar with the consequen t “annihilation” of the 5-7 pair into a 6-6 pair. The resulting capsid has the usu al morphological units, p enta mers and hexamers, but n ot th e spherical shap e. Thus it is to b e exp ected that in real viru ses scars s hould n ot b e visib le in the final stage but they should drive a change in shap e from spherical to non-spherical. It is plausible, though, that i) in exp erimen ts wh ere artificial virus capsids are synthesized, scars could b e actually seen at an in termediate stage of the assem bly wh en the “would-b e-capsid” is f r ozen at a su itable p oint in time; ii) n ot all scars are annihilated, hence s ome of th em could b e visible on the final capsid. Note that in the presence of scars, the total num b er of proteins needed is the s ame as for the icosadeltahedral case without scars (this fol- lo ws from 6 + 6 = 5 + 7) while the num b er and t yp e of morph ologica l units c hanges (for one simple scar: 13 p entamers, 1 h eptamer, 10 T − 12 hexamers, etc.). As said earlier, there is a strong interest to da y in try in g to generalize the CK construction to in clude non-sp herical viruses, imp ortant examples b eing the retrovi ru s es that ha v e spherical, sphero cylind rical and conical capsids (see, e.g., Ref.[14] and r eferen ces therein). In th e w ork of Ref.[6] th e prop osal that s p on taneous cu rv ature of the proteins in the capsids can driv e a change in shap e fr om sp herical to sph ero cylindrical or conical shap es is extensiv ely studied and th e geometric construction of certain capsids (sphero cilynd rical and conical) is carried out. T he application of that approac h to the case of retro viruses is then p erformed in Ref.[14], where the imp ortance of th e en vironment for the assembly of retro virus capsids is p ointed out. What w e conjecture here is that the b asis of these phenomena is the formation of scars. Our b elief is b ased on the follo win g observ ations: i) S cars app ear in the geometrica lly related (generalized) Thomson p roblem; ii) Their forma- tion/annihilation mechanism here seems to us a natural w a y to conv ert the energy giv en b y the en vironment into b ending energy (formation) and sub- sequen tly into stretc hing energy (annihilation); iii) This wa y a mec hanism for pro du cing a great v ariet y of shap es (not only the sphero cilyndrical or conical) is in place: the f ormation/annihilation of scars (simple or complex) in different lo cations on the intermediate icosadeltahedron (w e supp ose that this h as to happ en near the v ertices). Other authors h a v e sp eculated that scars should o ccur in viru s capsids [13]. T hey exp ect scars to b e formed only on large viru ses and that means 6 C 5 Scars:5-7-5 Pentamers Figure 3: The in termediate sph erical (icosadeltahedral) capsid with the C 5 - symmetric distribution of simp le scars. that they are exp ecting scars to b e seen on th e final capsid. This is an instance that w e d o not exclude but that is not essentia l for us as our main prop osal is to ascrib e the shap e c hange to the scars formation/annihilation mec hanism. V ariet y of shap es It is easy to con vince oneself that indeed a great v ariet y of shap es could b e obtained via th e scar form ation/annihilatio n mec hanism: A t the site on the intermediate icosadeltahedron where th e scar is formed and then annihilated th e sph ere gets stretc hed. The amount of s tretc h ing dep ends on the complexit y of the scar 5 . T he scars could b e formed symmet- rically (as w e sh all see in the next paragraph, for a particular s ymmetry of formation of scars we shall naturally obtain the sphero cylind rical shap e) or asymmetrically hence giving rise to r egular or irregular shap es. Of all these v ery large num b er of sh ap es only a su bset w ill d escrib e real virus capsids b ecause not all the shap es will b e stable or energetically fa vo red. A sys tem- atic study can b e carried on u s ing this metho d and case b y case it could b e seen wh ether it fits w ith the elastic prop erties of the virus capsids and with the constrain ts coming from the environmen t [14]. Wh at w e shall do no w is to constru ct, within our framework, one p articular shap e, the sphe- ro cylindrical. T his will giv e us the opp ortu n it y to sh o w how the m etho d of construction w orks in a case that it is kno wn to corresp ond to real v ir us capsids, like, e.g., certain bacteriophages. Sphero cylindrical Capsids Supp ose that the intermediate icosadelta- hedron is formed. W e can then r efer to the hexagonal lattice and to the template of Fig.1 and Fig.2. Let us imagine that the scars, e.g. all simple, are created only near the 10 inner v ertices via a mechanism th at r esp ects 5 Complex scars might not b e t hat rare as the same amount of energy is need ed for the formation of, sa y , one next-to-simple scar (5-7-5-7-5) and tw o simple scars, i.e. 4 E b . 7 Scarsare annihilated A A A A A A A A A A A B A A A A C C A A B C C Scarsare formed A A A A A 2 1 Figure 4: The generalized CK construction of the template dr iv en b y the scars form ation-annihilatio n mec h anism. the C 5 rotation symmetry 6 around the north p ole-south p ole axis 7 . I n Fig.3 the v ertices wh ere the scars are f ormed are indicated with • , while the other t w o are indicated with ◦ . T ak e a p air of the equilateral triangles of that template: an y one from one of the outer la y ers of fi v e triangles (e.g. the la y er of triangles that corresp ond to the north p ole) and the one from the inner la y er that sh ares an edge with it. In Fig.4 of such p airs is shown and the differen t n ature of the ve rtices is repr esen ted lik e in Fig.3. The scars are distributed in a wa y that is asymmetric with resp ect to the tw o triangles, hence the net effect of their formation/annihilation m echanism w ill deform them differentl y . Dep ending on the actual orien tation of the scar around the giv en v ertex the d eformation will b e different. T o obtain th e sph ero cylin- drical capsid the thr ee scars should mak e th e low er triangle thin ner and longer (they stretc h the area and mak e it bigger) and this h as the effect of shrinking the up p er triangle by m aking the common edge shorter. Due to the sym m etry of the lo cation of scars the tw o edges of th e new lo w er trian- gle ha v e to b e the s ame. If this mec hanism take s p lace in the same fashion for all the ten pairs 8 of triangles of the template of Fig.1 th e resulting new template is the one giv en in Fig.4. W e require that th is mec hanism is area preserving, i.e. that the total num b er of pr oteins needed is the same as the one needed for the icosadeltahedron, they are only rearranged. T his is obtained by requir in g that 2 α A = α 1 + α 2 , where α 1 is the area of the upp er new triangle and α 2 the area of the lo w er new triangle in Fig.4 . This means 6 C n is the fin ite group of rotations of angles 2 π /n , with n = 1 , 2 , 3 , ... . C 5 is one of the subgroups of the icosahedral group, the group of all p ossible symmetries of the icosahedron. I ts relev ance for the Thomson problem has b een understo od in [5] where a mec hanism of sp ontaneous symmetry breaking is seen as the resp onsible for some of the patterns found in numerical sim ulations. H ere our introdu ction of th e C 5 symmetry is motiv ated solely by t h e need to build up the sphero cylinder. 7 Of course the axis is completely arbitrary as long as it encompasses tw o opp osite vertices . 8 Five north p ole triangles paired with t heir common-edge inner triangles and five south p ole triangles paired with th eir common-ed ge inn er triangles. 8 B A A B B C C C C C B B B A A A A A A Figure 5: The sphero cylindr ical capsid. that the three quantiti es must b e related as √ 3 A 2 = B r A 2 − 1 4 B 2 + r C 2 − 1 4 B 2 ! , (4) with B < A and C > A . Recall that, for a = 1, A 2 = T = h 2 + k 2 + hk , hence th e final capsid, obtained by folding th e new template of Fig.4 (see Fig.5), will hav e (12 p entamers and ) th e 10( T − 1) hexamers d istributed differen tly with resp ect to th e inte rmediate icosadeltahedron. Notice that this sphero cylinder is s lightly d ifferen t from the one obtained in [6 ] as the u pp er and lo wer half-icosadelta hedr ons are n ot obtained b y folding fi ve equilateral triangles b ut five isosceles triangles (in this sense they are n o longer prop er h alf-icosa deltahedrons but a deformation of them). This is an instance that could b e exp erimentall y tested. F rom this construction it is clear that a v ariet y of shap es could b e ob - tained this wa y . F or instance, if the orient ation of the scars in the previous setting is such that C s h rinks, hen ce B b ecomes longer, th en a disk-lik e shap e is obtained. Let us stress h ere again that for th is to corresp ond to real viru s capsids one needs more d etailed information on the elastic prop- erties of th e proteins and of th e inte raction with the en vironment. 4 Conclusions In this pap er we prop ose a m echanism of formation and subsequent anni- hilation of scars of p en tamers-heptamers at an in termediate stage of the assem bly of the viru s capsid as the r esp onsible for a great v ariet y of non- spherical v ir us shap es. Our conjecture is based on the fact that scars are found in the (generalized) Th omson problem, in exp eriments and in numer- ical sim ulations, and on the observ ation that this mec h anism w ould giv e a simple and p lausible explanation of ho w the energy pro vided by the envi- ronment is conv erted in to a c hange of capsid’s shap e. The conjecture can b e tested, f or instance, in exp eriment s wh er e artificial capsids are synthesized. 9 Scars shou ld app ear on what w e called here the intermediate icosadeltahe- dron, th en should dr iv e the change in shap e. Caps ids that could p erhaps b e used to this en d are those relativ e to viruses that are kno wn to ha v e n on- spherical final shap e but still p en tamers and hexamers as morph ologic al units, like for instance certain b acteriophages. T his conjecture, if exp eri- men tally confirmed, w ould extend the classic Caspar and Klug construction for icosahedral viruses to include viruses that still h a v e p en tamers and hex- amers as morp hologica l u nits b ut no longer are icosadeltahedrons. Let us conclude by m aking the remark that a b etter und er s tanding of the w a y virus capsids are formed migh t suggest w a ys of destro ying a virus b y , for example, making th e capsid un stable. Ac kno wledgmen ts A.I. thanks Paul V o orheis of T rinit y College Dublin, Daniel Gru m iller of MIT Boston, for enj o y able discussions and for providing some difficult-to- find references and Ariann a Calistri of the Univ ersit y of Padua for advice with virology . S.S. ac kno wledges the kind hospitalit y of the Institute of P article and Nuclear Physics of Charles Univ ersit y Pr ague. References [1] W. Ch iu, R. M. Bur nett, R. L. Garcea Ed s., Structur al Biology of Viruses, Oxford Univ ersit y Press (New Y ork) 1997. [2] F. Crick, J. D. W atson, Nature 177 (1956) 473. [3] D. Caspar, A. Klug, Cold S pring Harb Symp. Quan t Biol. 27 (1962) 1. [4] Viru s Particle ExploreR (VIPER), V. Red d y ,P . Nata ra jan,B. Ok er- b erg,K. Li,K. Damo daran , R. Morton,C. Bro oks I I I , J. Johnson, J. of Vir. 75 (2001) 11943 (h ttp://vip erdb.scrip p s.edu/) [5] A. Iorio, S. Sen, Phys. Rev. B 74 (2006 ) 052102; ibid em 75 (200 7) 099901 (E). [6] T. T. Nguy en, R. F. Bruinsma, W. M. Gelbart, Phys. Rev. E 72 (2005 ) 05192 3. [7] J. Lid m ar, L. Mirn y , D. R. Nelson, Ph ys. Rev. E 68 (2003) 05191 0. [8] J. J. Thomson, Phil. Mag. 7 (1904) 237. [9] M. J . Bo wic k, D. R. Nelson, A. T ra v esset, Ph ys. Rev. B 62 (2000) 8738. [10] M. J . Bo wic k, A. C acciuto, D. R. Nelson, A. T r a v esset, Phys. Rev. Lett. 89 (200 2) 185 502 and Phys. Rev. B 73 (2006) 024115 . 10 [11] E . L. Altsc huler, T. J. Williams, E. R. Ratner, R. Tipton, R. Stong, F. Do wla, F. W o oten, Prh ys. Rev. Lett. 78 (1997) 2681. [12] A. Pe rez-Garrido, M. J . W. Do dgson, M A. Mo ore, M. Ortu ˜ no, A. Diaz- Sanc hez, Phys. Rev. Lett. 79 (1997) 1417. [13] A. R. Bausc h, M. J. Bo wic k, A. Cacciuto, A. D. Dinsmore, M. F. Hsu, D. R. Nelson, M. G. Nik olaides, A. T ra v esset, D. A. W eitz, Science 299 (2003 ) 1716 . [14] T . T. Nguy en, R. F. Bruinsm a, W. .M. Gelbart, Ph ys. Rev. Lett. 96 (2006 ) 0781 02. 11
Original Paper
Loading high-quality paper...
Comments & Academic Discussion
Loading comments...
Leave a Comment